Potrebbero piacerti anche
- Elementos Basicos de CitologiaDocumento12 pagineElementos Basicos de CitologiaKakb Estudios100% (1)
- Significado Psicoanalitico Tatuajes y PiercingsDocumento2 pagineSignificado Psicoanalitico Tatuajes y PiercingsDresan PezloNessuna valutazione finora
- Denticion PrenatalDocumento45 pagineDenticion PrenatalJosephDelgadoDelgado0% (2)
- Trauma de TóraxDocumento31 pagineTrauma de TóraxPamela Bahamondes Manosalva100% (1)
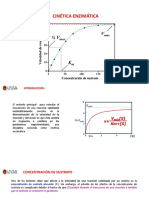
- Tema 8 - Cinetica Enzimatica Farmacia PDFDocumento12 pagineTema 8 - Cinetica Enzimatica Farmacia PDFMariciell Larissa Gonzales TiconaNessuna valutazione finora
- ECOEDocumento4 pagineECOEKarlos Asto Altamirano100% (3)
- 2022 Manual Manejo ClinicoDocumento221 pagine2022 Manual Manejo ClinicoPriscilla PaivaNessuna valutazione finora
- Tubo digestivo IIDocumento9 pagineTubo digestivo IIRegina Cortés100% (1)
- PH PracticaDocumento9 paginePH PracticaeuclidesraulNessuna valutazione finora
- Aspiración de Secreciones Con Circuito Cerrado y Manejo de Dispositivos en Via AereaDocumento42 pagineAspiración de Secreciones Con Circuito Cerrado y Manejo de Dispositivos en Via Aerearepre6467% (3)
- Counselling Con Adolescentes Mejor Con La FamiliaDocumento122 pagineCounselling Con Adolescentes Mejor Con La Familiajeraldinegaray100% (1)
- AntihemorrágicosDocumento21 pagineAntihemorrágicoskprh311100% (7)
- Fármacos cardiológicos: inotrópicos, cronotrópicos, dromotrópicos y batmotrópicosDocumento2 pagineFármacos cardiológicos: inotrópicos, cronotrópicos, dromotrópicos y batmotrópicosalexander puma sacsahuillca100% (2)
- Técnica Patch ClampDocumento3 pagineTécnica Patch ClampAlejandroHidalgoNavaNessuna valutazione finora
- Neurociencias Clase I Neuronas Circuitos Neuronales y ConductaDocumento19 pagineNeurociencias Clase I Neuronas Circuitos Neuronales y ConductaMacarena CepedaNessuna valutazione finora
- Resumen Neurobiología FINALDocumento98 pagineResumen Neurobiología FINALCynthia ScottiNessuna valutazione finora
- Permeabilidad de La Membrana CelularDocumento5 paginePermeabilidad de La Membrana CelularCamila ContrerasNessuna valutazione finora
- Presentacion 6 Cinetica EnzimaticaDocumento46 paginePresentacion 6 Cinetica EnzimaticaE Clinton LMNessuna valutazione finora
- Trabajo Potencia y Energia Aplicado A La SaludDocumento24 pagineTrabajo Potencia y Energia Aplicado A La Saludashli trujillo0% (1)
- Gradiente ElectroquímicoDocumento3 pagineGradiente ElectroquímicoGeralNessuna valutazione finora
- Metabolismo Intermediario y AnabolismoDocumento8 pagineMetabolismo Intermediario y AnabolismoAmyEBermudezQuero0% (1)
- La Célula AnimalDocumento4 pagineLa Célula AnimalSERGIO MENA PEREZ ESTUDIANTE ACTIVONessuna valutazione finora
- Gradiente electroquímico y fuerza protón-motrizDocumento4 pagineGradiente electroquímico y fuerza protón-motrizRenzzo B NamucheNessuna valutazione finora
- Informe de Aminoácidos - Grupo 9Documento9 pagineInforme de Aminoácidos - Grupo 9Jonathan Andres Ortiz ForeroNessuna valutazione finora
- Ejercicio 1 EnzimologiaDocumento15 pagineEjercicio 1 EnzimologiaAndrea ChavarroNessuna valutazione finora
- Fuerza Iónica y CoeficienteDocumento12 pagineFuerza Iónica y CoeficienteMaria Angelica Oviedo TorresNessuna valutazione finora
- Fermentacion 3Documento10 pagineFermentacion 3rocioNessuna valutazione finora
- Informe EquilibrioDocumento13 pagineInforme EquilibrioEstefany Duran LondoñoNessuna valutazione finora
- Contracción Del Musculo Liso - FisiologíaDocumento5 pagineContracción Del Musculo Liso - FisiologíaDara DosNessuna valutazione finora
- Aspectos Biologicos de Gracilaria ChilensisDocumento5 pagineAspectos Biologicos de Gracilaria Chilensisemilymr3Nessuna valutazione finora
- Taller Señalización CelularDocumento5 pagineTaller Señalización CelularROXY FERNANDA RODRÍGUEZ TORRESNessuna valutazione finora
- Examen 3 MicrobiologiaDocumento41 pagineExamen 3 Microbiologiaapi-3806615100% (1)
- Purificación y caracterización de una lectina de habas mediante cromatografía y ensayos de actividad hemaglutinanteDocumento8 paginePurificación y caracterización de una lectina de habas mediante cromatografía y ensayos de actividad hemaglutinanteCesarNessuna valutazione finora
- Fundamentos de La TermodinamicaDocumento6 pagineFundamentos de La TermodinamicaAriane A Martinez RNessuna valutazione finora
- Informe 3 - Actividad EnzimáticaDocumento12 pagineInforme 3 - Actividad EnzimáticaAndrea SalinasNessuna valutazione finora
- Enzimas ResumenDocumento8 pagineEnzimas ResumenJuán Gabriel Gallego LópezNessuna valutazione finora
- GlucosaDocumento5 pagineGlucosalupemargaritaNessuna valutazione finora
- Pro ProteinasDocumento1 paginaPro ProteinasSosimo BenitezNessuna valutazione finora
- Niveles organización SN aprendizaje AplysiaDocumento3 pagineNiveles organización SN aprendizaje AplysiaAndreBelenNessuna valutazione finora
- Practica 3 Corregida Utp Glucolisis AnaerobiaDocumento6 paginePractica 3 Corregida Utp Glucolisis AnaerobiaJessica Valentina Zarama BuriticaNessuna valutazione finora
- La Molécula IntermediariaDocumento3 pagineLa Molécula IntermediariaGarcia Jonnattan0% (1)
- Metabolismo de CarboidratosDocumento23 pagineMetabolismo de CarboidratosVerónica VelásquezNessuna valutazione finora
- EnzimasDocumento23 pagineEnzimasMaruby MontonerNessuna valutazione finora
- Fisica BioelectricidadDocumento18 pagineFisica BioelectricidadKaren Ramirez100% (1)
- Autoevaluación 2.1 - Respiración Celular Biología UesDocumento3 pagineAutoevaluación 2.1 - Respiración Celular Biología UesMayra Marroquin50% (2)
- La EntalpíaDocumento6 pagineLa EntalpíaStefaniaNessuna valutazione finora
- Factores Que Inhiben La MitosisDocumento3 pagineFactores Que Inhiben La MitosisDavis AvilbarNessuna valutazione finora
- Mecanismos de Regulación EnzimáticaDocumento2 pagineMecanismos de Regulación EnzimáticaRobert Isaias Mendoza CardenasNessuna valutazione finora
- Guia 5. Actividad Enzimática UreasaDocumento8 pagineGuia 5. Actividad Enzimática UreasaAnderson Murcia PolaniaNessuna valutazione finora
- Tema 4. Tejido Parenquimático y MecánicoDocumento24 pagineTema 4. Tejido Parenquimático y MecánicoChristopher SilvaNessuna valutazione finora
- Transpiracion 06 PDFDocumento9 pagineTranspiracion 06 PDFJeanKeinerWyLopezNessuna valutazione finora
- Reporte p.4 FisioDocumento13 pagineReporte p.4 FisioSeccion 1, Reyes Sánchez Zuleima GuadalupeNessuna valutazione finora
- OriginalDocumento42 pagineOriginalCarlenis RamosNessuna valutazione finora
- Tema 2 - PROTEÍNASDocumento39 pagineTema 2 - PROTEÍNASJose Esteban De Haro MartinezNessuna valutazione finora
- Blanco. Cap 3. ProteinasDocumento36 pagineBlanco. Cap 3. ProteinasAldo CruzNessuna valutazione finora
- Cambios térmicos de un sistema: primeras leyes de la termodinámicaDocumento14 pagineCambios térmicos de un sistema: primeras leyes de la termodinámicablink42j0% (1)
- Tarea Escrita - Cruzamiento PruebaDocumento1 paginaTarea Escrita - Cruzamiento PruebaAna Victoria Ramírez IriarteNessuna valutazione finora
- Clase 4 - Regulación Actividad EnzimáticaDocumento7 pagineClase 4 - Regulación Actividad EnzimáticaanaNessuna valutazione finora
- Tabla Procesos MetabolicosDocumento10 pagineTabla Procesos MetabolicosValeria EnriquezNessuna valutazione finora
- Taller ProteínasDocumento5 pagineTaller ProteínasDiego RodriguezNessuna valutazione finora
- Fenómenos InterfacialesDocumento28 pagineFenómenos InterfacialesFernando100% (1)
- Manual FQF Practicas de LaboratorioDocumento78 pagineManual FQF Practicas de LaboratorioLuis Huante de PerafanNessuna valutazione finora
- Termogenesis y ObesidadDocumento12 pagineTermogenesis y ObesidadGerald ToroNessuna valutazione finora
- Sistema Nervioso de Los MamíferosDocumento5 pagineSistema Nervioso de Los MamíferosEliana PérezNessuna valutazione finora
- Señalización celular: tipos de señales y receptores enDocumento22 pagineSeñalización celular: tipos de señales y receptores enAndrésNessuna valutazione finora
- TEMA 47 Elongación Ácidos GrasosDocumento3 pagineTEMA 47 Elongación Ácidos GrasosAkimita88% (8)
- Potencial de Difusion y Potencial de LesionDocumento13 paginePotencial de Difusion y Potencial de LesionLatika CzNessuna valutazione finora
- Citoesqueleto-ActinaMicrotúbulosFilamentosDocumento46 pagineCitoesqueleto-ActinaMicrotúbulosFilamentosLeslie Zavaleta Bazan0% (1)
- Practica # 1de Botánica: Células VegetalesDocumento4 paginePractica # 1de Botánica: Células VegetalesBorreSugar100% (2)
- Patch ClampDocumento6 paginePatch ClampDaniela MonroyNessuna valutazione finora
- Complejo Del Poro NuclearDocumento11 pagineComplejo Del Poro NuclearElvisRojas972010Nessuna valutazione finora
- Quistes odontogénicos: clasificación, características y tratamientoDocumento7 pagineQuistes odontogénicos: clasificación, características y tratamientoGabyAvillasanteNessuna valutazione finora
- Disfunciones Del Aparato LocomotorDocumento7 pagineDisfunciones Del Aparato LocomotorCarlos CardonaNessuna valutazione finora
- Evaluacion Recuperatoria Piel AnimalDocumento2 pagineEvaluacion Recuperatoria Piel AnimalLina Luz Perez MedranoNessuna valutazione finora
- Enfermedades-Respiratorias Medicina 3Documento37 pagineEnfermedades-Respiratorias Medicina 3Ann' KateeNessuna valutazione finora
- Auscultación AngheloDocumento30 pagineAuscultación AngheloAnghelo Figueroa BustamanteNessuna valutazione finora
- Nutricion para DeportistasDocumento64 pagineNutricion para DeportistasValeria Duque GomezNessuna valutazione finora
- Alimentos de Origen Anima1Documento2 pagineAlimentos de Origen Anima1Jhonatan Vilela TinedoNessuna valutazione finora
- Taller de MusculosDocumento12 pagineTaller de MusculoslinaNessuna valutazione finora
- Sistema Respiratorio AvesDocumento1 paginaSistema Respiratorio Aves- r e d s u n -Nessuna valutazione finora
- Mary Bella AlavDocumento3 pagineMary Bella AlavoscarNessuna valutazione finora
- Apoptosis - Dra. Rina ZumaetaDocumento18 pagineApoptosis - Dra. Rina ZumaetaJuan VasquezNessuna valutazione finora
- La Fisiología Del AdultoDocumento3 pagineLa Fisiología Del AdultoMayhuiri FredNessuna valutazione finora
- 2° S.Nervioso 4 AnatomiaDocumento1 pagina2° S.Nervioso 4 Anatomiafabian alejandro santana leivaNessuna valutazione finora
- Informe 1Documento6 pagineInforme 1maugarage01Nessuna valutazione finora
- Transfusiones Sanguíneas EnfermeríaDocumento68 pagineTransfusiones Sanguíneas EnfermeríaJohnCardona50% (2)
- PPR Informwe FinalDocumento68 paginePPR Informwe FinalWilly Cabana PizarroNessuna valutazione finora
- Guía Mamíferos 2023Documento11 pagineGuía Mamíferos 2023leapopNessuna valutazione finora
- Diferenciacion Celular BiologiaDocumento12 pagineDiferenciacion Celular BiologiaMegevi GeldresNessuna valutazione finora
- 211 EBAU2020-JULIO-BIOLOGIA Examen y CriteriosDocumento6 pagine211 EBAU2020-JULIO-BIOLOGIA Examen y CriteriosSergio PootNessuna valutazione finora