Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Estructura de Los Tejidos Periodontales
Caricato da
Ariel Gottlieb Palombo100%(1)Il 100% ha trovato utile questo documento (1 voto)
106 visualizzazioni18 pagineCopyright
© © All Rights Reserved
Formati disponibili
PDF, TXT o leggi online da Scribd
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
© All Rights Reserved
Formati disponibili
Scarica in formato PDF, TXT o leggi online su Scribd
100%(1)Il 100% ha trovato utile questo documento (1 voto)
106 visualizzazioni18 pagineEstructura de Los Tejidos Periodontales
Caricato da
Ariel Gottlieb PalomboCopyright:
© All Rights Reserved
Formati disponibili
Scarica in formato PDF, TXT o leggi online su Scribd
Sei sulla pagina 1di 18
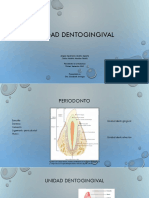
El periodoncio, definido como aquellos tejidos que
sostienen y revisten los dientes, est constituido por
el cemento radicular, el ligamento periodontal, el
hueso que reviste el alvolo dental (hueso alveolar)
y la parte de la enca que se une al diente (unin den-
togingival). La gran prevalencia de enfermedades pe-
riodontales y la comprensin de que los tejidos per-
didos pueden ser reparados y, quizs, regenerados ha
despertado un inters considerable en los factores
y las clulas que regulan la formacin y el manteni-
miento del tejido. Es importante entender que cada
uno de los componentes periodontales tiene su pro-
pia estructura especializada, y que estas caractersti-
cas estructurales definen directamente la funcin. De
hecho, el propio funcionamiento del periodoncio slo
se logra a travs de la integracin estructural y la in-
teraccin entre sus componentes.
En los aos recientes se han publicado numerosas
descripciones detalladas de las caractersticas estruc-
turales y de composicin de los tejidos periodontales
(3, 5-7, 9, 15, 17, 46, 50, 56, 58, 61); se remite al lector
a esta bibliografa para una descripcin ms amplia
del desarrollo, la formacin y la estructura de los teji-
dos periodontales. Esta revisin se centrar en las re-
laciones estructura-funcin necesarias para compren-
der la destruccin tisular periodontal y la reparacin
o regeneracin de las estructuras afectadas.
Tejidos periodontales sanos
La unin dentogingival
La unin dentogingival (la enca que se une al diente)
es una adaptacin de la mucosa bucal que comprende
componentes tisulares epiteliales y conectivos. El epi-
telio se divide en tres compartimientos funcionales
epitelio gingival, sulcular y de unin, y el tejido co-
nectivo, en los compartimientos superficial y pro-
fundo. El epitelio de unin desempea una funcin
crucial, ya que esencialmente asla los tejidos perio-
dontales del entorno bucal. Su integridad es, por lo
tanto, esencial para mantener un periodoncio sano.
La enfermedad periodontal se instaura cuando la es-
tructura del epitelio de unin empieza a fallar, lo que
constituye un excelente ejemplo de cmo la estruc-
tura determina la funcin.
El epitelio de unin
El epitelio de unin surge del reducido epitelio del
esmalte a medida que el diente erupciona dentro de la
cavidad bucal. Forma un collar alrededor de la porcin
cervical del diente a continuacin de la unin amelo-
cementaria (fig. 1). La superficie libre de este collar
constituye el suelo del surco gingival. Bsicamente, el
epitelio de unin es un epitelio indiferenciado, esca-
moso, estratificado, con una elevada frecuencia de re-
cambio celular. Es ms grueso cerca del fondo del surco
gingival, y su grosor disminuye a medida que desciende
apicalmente a lo largo de la superficie dentaria, hasta
alcanzar el grosor de unas pocas clulas. Este epitelio
est constituido por clulas aplastadas, paralelas al
diente, que derivan de una capa de clulas basales cu-
boides situada fuera de la superficie dentaria y que des-
cansa sobre una membrana basal. Las clulas supra-
basales tienen una ultraestructura similar y, de forma
bastante notable, mantienen la capacidad de experi-
mentar divisin celular. La capa de clulas que se une
al diente proporciona la insercin real de la enca a la
superficie dentaria mediante un complejo estructural
denominado la insercin epitelial. Este complejo consta
de una estructura tipo lmina basal que es adherente
a la superficie del diente y a la que la capa de clulas
superficiales adhiere por hemidesmosomas. La estruc-
tura tipo lmina basal es una matriz extracelular espe-
11
Periodontology 2000 (Ed Esp), Vol. 16, 2007, 11-28
Estructura de los tejidos periodontales
en el individuo sano y en el individuo
enfermo*
ANTONIO NANCI Y DIETER D. BOSSHARDT
Copyright Blackwell Munksgaard
PERIODONTOLOGY 2000
ISSN 0906-6713
Copyright Grupo Ars XXI de Comunicacin, S.L.
PERIODONTOLOGY 2000 (Ed Esp)
ISSN 1695-1808
*Parte de este artculo est adaptado de la cita bibliogrfica 30.
cializada en la que no han sido inmunodetectados los
constituyentes tpicos de la membrana basal en una
cantidad significativa, pero que es rica en glucoconju-
gados y contiene laminina 5. Esta protena de la ma-
triz es mediadora de la adhesin celular y regula la po-
larizacin y la migracin de los queratinocitos (27).
Las clulas del epitelio de unin difieren conside-
rablemente de las del epitelio gingival. Contienen ms
citoplasma, un retculo endoplsmico rugoso y cuer-
pos de Golgi. Exhiben menos tonofilamentos y des-
mosomas, as como espacios intercelulares ms an-
chos. Estos espacios, llenos de lquido, normalmente
contienen leucocitos polimorfonucleares y monocitos
que pasan desde el tejido conectivo subepitelial a surco
gingival, a travs del epitelio de unin. Las clulas mo-
nonucleares, juntamente con las molculas segrega-
das por ellas y otras originadas por los fluidos de las
clulas del epitelio de unin, la sangre y los tejidos,
representan la primera lnea de defensa en el control
del perpetuo desafo microbiano. Entre estas molcu-
las se encuentran las -defensinas, las -defensinas,
la catelicidina IL-37, las interleucinas (IL) IL-8, IL-1
e IL-1, el factor de necrosis tumoral (TNF-), la
molcula de adhesin intercelular tipo 1 y el antgeno
tipo 3 asociado a la funcin linfocitaria.
El compartimiento de tejido conectivo
El tejido conectivo que sostiene al epitelio de unin
es estructuralmente diferente del que sostiene al epi-
telio gingival bucal. Incluso en circunstancias clnica-
mente normales, muestra un infiltrado celular infla-
matorio. El tejido conectivo adyacente al epitelio de
unin contiene un plexo vascular extenso. Las clulas
inflamatorias, tales como los leucocitos polimorfonu-
cleares y los linfocitos T, se extravasan continuamente
de este denso entramado de capilares y vnulas pos-
capilares y migran a travs del epitelio de unin hacia
el surco gingival y, finalmente, hacia el fluido bucal. En
Schroeder y Listgarten (58) se describe en detalle la
distribucin vascular en la lmina propia gingival.
El epitelio de unin puede ser considerado un epi-
telio escamoso estratificado desarrollado de forma in-
completa. Alternativamente, tambin es visto como
una estructura que evoluciona a lo largo de una tra-
yectoria diferente y produce los componentes de la in-
sercin epitelial en vez de progresar ms hacia un epi-
telio queratinizado. Se cree que la especial naturaleza
del epitelio de unin refleja el hecho de que el tejido
conectivo que lo sostiene es funcionalmente diferente
del que sostiene al epitelio sulcular, una diferencia con
importantes implicaciones para entender el avance de
la enfermedad periodontal y la regeneracin de la unin
dentogingival despus de la ciruga periodontal. Se cree
que el tejido conectivo subepitelial (lmina propia)
proporciona seales instructivas para la progresin
normal del epitelio escamoso estratificado (36, 38). Esta
sealizacin supuestamente es inexistente en los teji-
dos conectivos ms profundos, de modo que el epite-
lio en contacto con stos no alcanza el mismo grado
de diferenciacin.
Por consiguiente, el epitelio sulcular, a diferencia
del epitelio gingival, no es queratinizado, aunque am-
bos son sostenidos tcnicamente por una lmina pro-
pia similar. Esta diferencia en la expresin epitelial
tambin puede ser atribuida a la inflamacin. Incluso
bajo condiciones clnicas normales, el tejido conec-
tivo asociado con la unin dentogingival est ligera-
mente inflamado. En animales experimentales se ha
comprobado que si el proceso inflamatorio se elimina,
implementando un estricto rgimen de higiene bu-
codental combinado con cobertura antibitica, el epi-
telio sulcular queratiniza (21, 22).
El cemento
El cemento es el tejido conectivo duro, avascular,
que recubre las races de los dientes y que sirve prin-
Nanci y Bosshardt
12
Fig. 1. Microfotografa electrnica de retrodispersin de una
seccin de tejido descalcificada que muestra la regin cer-
vical de un diente de rata con el epitelio de unin (EU), el
espacio del esmalte (EE) y la unin amelocementaria (UAC).
Se observan numerosos vasos sanguneos (VS) en el tejido
conectivo (TC) de la lmina propia. Ntese como el espacio
del esmalte se extiende entre el cemento y la dentina (ca-
beza de flecha), una situacin que puede dar la impresin
de que existe una capa intermedia entre ellos. D: dentina.
EU
VS
D
TC
A
UAC
50 m
EE
cipalmente para revestir y adherir las principales fi-
bras del ligamento periodontal. Bsicamente, existen
dos variedades de cemento distinguidas en funcin
de la presencia o la ausencia de clulas en su interior
y el origen de las fibras de colgeno de la matriz.
Variedades de cemento
El cemento acelular de fibras extrnsecas (acellular
extrinsic fiber cementum, AEFC) (cemento primario o
cemento acelular) se encuentra desde la mitad cervi-
cal hasta dos tercios de la raz (figs. 2-4). Se desarrolla
muy lentamente, y se considera que es acelular por-
que las clulas que lo forman permanecen sobre su su-
perficie. El nmero tan elevado de fibras de colgeno
del ligamento periodontal que se insertan en el AEFC,
donde se las denomina fibras de Sharpey, seala su
importante funcin en la adherencia dentaria. El grado
general de mineralizacin del AEFC es de alrededor del
45-60 %, pero el examen radiogrfico con poca radia-
cin revela que la capa ms interna est menos mine-
ralizada y que las capas ms externas estn caracteri-
zadas por bandas alternadas, con mayor o menor
contenido mineral, paralelas a la superficie de la raz.
El cemento celular de fibras intrnsecas (cellular in-
trinsic fiber cementum, CIFC) (cemento secundario
o cemento celular) est distribuido a lo largo del ter-
cio apical o la mitad apical de la raz y en las reas de
furcacin (fig. 5). Puesto que el cemento celular de fi-
bras intrnsecas tambin es producido como un tejido
de reparacin que rellena los defectos de resorcin y
las fracturas radiculares, tambin puede encontrarse
ms coronalmente. El colgeno producido por los ce-
mentoblastos (fibras de colgeno intrnsecas) y la pre-
sencia de cementoblastos atrapados en lagunas den-
tro de la matriz que ellos producen (cementocitos) son
los rasgos caractersticos del cemento celular de fibras
intrnsecas. La heterognea organizacin del colgeno,
su rpida velocidad de formacin y la presencia de c-
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
13
Fig. 2. Microfotografa electrnica de retrodispersin que
muestra el desarrollo del cemento de fibra extrnseca ace-
lular (AEFC) en un premolar humano desde la porcin api-
cal (A) hasta la cervical (B, C). A). Tras la desintegracin de
la vaina radicular epitelial de Hertwig (HERS), las clulas
que estn en la superficie de la raz expuesta implantan una
franja de fibras colgenas (FF) en la matriz de dentina an
sin mineralizar (PD: predentina). B) La franja de fibras est
orientada perpendicularmente a la superficie radicular y
envuelve las clulas adyacentes. C) Cuando la capa de ce-
mento alcanza un grosor de aproximadamente 10 m, la
mayora de las fibras de la franja todava son cortas, mien-
tras que otras se han alargado hasta el espacio del liga-
mento periodontal (LP). D: dentina; FM: frente de minera-
lizacin; P: pulpa; Od: odontoblastos.
D
Od
PD
P
FF
LP
D
D
AEFC
FM
HERS
50 m 10 m 10 m
A B C
lulas y lagunas pueden explicar porque esta variedad
de cemento se encuentra menos mineralizada que el
cemento acelular de fibras extrnsecas.
El cemento celular de fibras intrnsecas constituye el
componente intrnseco del cemento celular estratificado
mixto (cellular mixed stratified cementum), que posee
Nanci y Bosshardt
14
Fig. 3. Microfotografa electrnica de transmisin que ilus-
tra la superficie radicular cervical de un diente humano. El
cemento acelular de fibras extrnsecas (AEFC), que preva-
lece en esta regin de la raz, est caracterizado por fibras
del ligamento periodontal (FLP) densamente apretadas,que
entran en la capa de cemento a travs del frente de mine-
ralizacin (FM). Cb: cementoblastos; N: ncleo.
FLP
Cb
N
FM
AEFC
1 m
A
Fig. 4. Microfotografa electrnica del cemento acelular de fi-
bras extrnsecas (AEFC) de secciones tisulares, tras el inmu-
nomarcado con oro para la sialoprotena sea (BSP). A) Las
fibras del ligamento periodontal (FLP) que entran en la capa
de cemento a travs del frente de mineralizacin (FM). El mar-
cado predomina sobre la matriz de cemento interfibrilar. B)
En la regin de la unin dentina-cemento, las fibrillas de
colgeno cementarias y dentinales se superponen y se en-
trecruzan. Las protenas no colgenas, como la sialoprotena
sea, rellenan los anchos espacios interfibrilares. D: dentina.
A
B
BSP
FLP
FM
AEFC
AEFC
D
500 m 1 m
una estratificacin derivada del depsito consecutivo de
capas alternadas de cemento acelular de fibras extrn-
secas y de cemento celular de fibras intrnsecas. El ce-
mento celular estratificado mixto no se encuentra en los
molares de los roedores, pero siempre est presente en
los dientes humanos. Puesto que la variedad de cemento
intrnseco puede formarse focalmente, y con mucha ra-
pidez, puede servir como un medio para ajustar la po-
sicin dentaria a nuevos requerimientos.
Composicin bioqumica del cemento
La composicin del cemento se parece a la del hueso.
Contiene una matriz que es aproximadamente un 50 %
mineral (apatita substituida) y un 50 % orgnica. El co-
lgeno tipo I es el componente orgnico predominante,
constituyendo hasta el 90 % de la matriz orgnica. En-
tre los otros colgenos asociados con el cemento se in-
cluyen el colgeno tipo III, un colgeno menos entre-
cruzado que se encuentra en altas concentraciones
durante el desarrollo y reparacin o regeneracin de los
tejidos mineralizados, y el colgeno tipo XII, un col-
geno asociado a la fibrilla con triples hlices interrum-
pidas (fibril-associated collagen with interrupted triple
helices, FACIT) que se une al colgeno de tipo I y tam-
bin a las protenas no colgenas de la matriz. En ex-
tractos de cemento maduro tambin es posible hallar
escasas cantidades de otros colgenos, como los de tipo
V, VI y XIV; sin embargo, stos pueden ser contaminan-
tes procedentes de la regin del ligamento periodontal
asociada con las fibras insertadas en el cemento. Casi
todas las protenas no colgenas de la matriz identifi-
cadas en el cemento tambin se encuentran en el hueso
(7): sialoprotena sea (BSP) (fig. 4), protena 1 de la
matriz de la dentina (DMP-1) (20, 24, 44), sialoprotena
dentinaria (1), fibronectina, osteocalcina, osteonectina,
osteopontina, tenascina (47, 69), proteoglucanos, pro-
teolpidos y varios factores de crecimiento, entre los que
se incluyen el factor de crecimiento del cemento, que
parece ser una molcula tipo factor de crecimiento in-
sulinoide (IGF). Asimismo, se ha sugerido la presencia
en el cemento de protenas del esmalte. Se ha infor-
mado que las clulas de la vaina radicular epitelial de
Hertwig (HERS) pueden sintetizar las amelogeninas que
se acumulan en la superficie de la raz que se est for-
mando, constituyendo una capa denominada cemento
intermedio (40-42, 60). Hasta la fecha, sin embargo, no
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
15
Fig. 5. Microfotografa electrnica de retrodispersin que
muestra el desarrollo del cemento de fibra intrnseca celu-
lar (CIFC) en un premolar humano desde direccin apical
(A) hasta direccin coronal (B, C). A) Tras la desintegracin
de la vaina radicular epitelial de Hertwig (HERS), los ce-
mentoblastos (Cb) en la superficie radicular expuesta de-
positan rpidamente la matriz de cemento sobre la matriz
de dentina todava sin mineralizar (PD: predentina). B) Al-
gunos cementoblastos se incrustan como cementocitos (Cc)
en su propia matriz. C) En una capa de cemento ms ma-
dura y gruesa, los cementocitos se alojan en lagunas. D,
dentina; Od, odontoblastos; PGE
2
, prostaglandina E
2
.
CIFC
D
Cc
Cb
PD
D
CIFC
CIFC
Cb
Cc
Cc
D
10 m
Od
HERS
A B C
se dispone de pruebas conclusivas con respecto a que
las amelogeninas o las no amelogeninas se acumulen
en los constituyentes de la matriz del cemento normal
o incluso que formen una capa distinta entre la dentina
y el cemento. Aunque se encuentran protenas de la ma-
triz del esmalte sobre la raz, su presencia est limitada
a una regin muy corta, cervical, que probablemente
representa la extremidad cervical de la corona sobre la
cual se deposita el cemento (11, 12). Tambin se ha co-
municado la espordica expresin de protenas del es-
malte a lo largo de la raz en los dientes porcinos (fig.
6) (13) y en los molares de roedores en asociacin con
las clulas epiteliales atrapadas en el cemento celular
de fibras intrnsecas (12, 25, 26, 63). Finalmente, tam-
bin se ha identificado en el cemento una protena de
adhesin al cemento aparentemente diferente (64).
Desarrollo del cemento
La formacin del cemento tiene lugar a lo largo de
toda la raz y durante toda la vida del diente. Sin em-
bargo, su inicio est limitado al avance del borde radi-
cular durante la formacin de la raz. Se cree que en esta
zona la vaina radicular epitelial de Hertwig, que deriva
de la extensin apical del epitelio interno y externo del
esmalte, enva un mensaje inductivo, posiblemente me-
diante la secrecin de determinadas protenas de la
matriz del esmalte, a las clulas pulpares ectomesen-
quimatosas opuestas. Estas clulas se diferencian en
odontoblastos y producen una capa de predentina.
Poco despus, la HERS se fragmenta, y las clulas ec-
tomesenquimatosas de la porcin interna del folculo
dentario pueden entonces entrar en contacto con la
predentina. Algunas clulas de la vaina radicular frag-
mentada forman grupos separados, rodeados por una
membrana basal, conocidos como los restos epitelia-
les de Malassez, que persisten en el ligamento perio-
dontal maduro. Posteriormente, los cementoblastos se
diferencian y depositan la matriz de cemento sobre la
dentina radicular en formacin. No estn totalmente
esclarecidos el origen de los cementoblastos y la serie
de acontecimientos que culminan en su diferencia-
cin, y estos temas se analizarn a continuacin.
Formacin del cemento
Cemento acelular de fibras extrnsecas: durante el de-
sarrollo de la raz en los dientes humanos, las prime-
Nanci y Bosshardt
16
Fig. 6. Microfotografa electrnica de transmisin que mues-
tra la superficie cervical de la raz, al comienzo del proceso de
formacin de la raz en diente porcino; la seccin tisular fue
procesada con el anticuerpo antiamelogenina, mediante in-
munomarcado con oro. A lo largo de la superficie de dentina
se observan, espordicamente, masas de matriz que contie-
nen amelogenina (cabezas de flecha), junto con la matriz co-
lgena de precemento (PC). AMEL: amelogenina; N: ncleo.
AMEL
N
PC
A
D
500 nm
ras clulas que se alinean a lo largo de la superficie del
recin formado manto de dentina, todava sin mine-
ralizar, exhiben caractersticas fibroblsticas (fig. 2). Es-
tas clulas depositan colgeno dentro de la matriz de
la dentina no mineralizada, de modo que las fibrillas
de ambas matrices se entrelazan. La mineralizacin
del manto de dentina se inicia de forma interna y no
llega a la superficie hasta que las fibrillas de colgeno
de ambas capas se combinan. Luego se dispersa hacia
el interior de la matriz de cemento, estableciendo as
la unin dentina-cemento. El cemento acelular de fi-
bras extrnsecas inicial, por lo tanto, consiste en una
fina capa mineralizada con una corta franja de fibras
de colgeno implantadas perpendiculares a la super-
ficie de la raz. Las clulas que se encuentran en la su-
perficie radicular continan depositando colgenos,
de forma que la corta franja se alarga y su grosor au-
menta. Al mismo tiempo, tambin secretan protenas
no colgenas de la matriz, que rellenan los espacios
entre las fibras de colgeno y regulan la mineraliza-
cin de la capa de cemento que se est formando (fig.
4). Esta actividad contina hasta que se queda consti-
tuida una capa de cemento de aproximadamente 15-
20 m, momento en el cual la franja de fibras intrn-
secas se conecta al haz de fibras del ligamento
periodontal en desarrollo (fig. 3). Posteriormente, las
clulas que forman el cemento acelular de fibras ex-
trnsecas estarn esencialmente ocupadas en la snte-
sis de las protenas no colgenas de la matriz; las fi-
brillas de colgeno que se incrustan en l sern
formadas por los fibroblastos del ligamento perio-
dontal. No existe en la superficie del cemento acelu-
lar de fibras extrnsecas una capa morfolgicamente
distinguible de cementoide, semejante al osteoide o a
la predentina. Aunque esta variedad de cemento est
clasificada como cemento de fibras extrnsecas, podra
tal vez afirmarse que en el inicio del proceso es un ce-
mento de fibras intrnsecas. Tal como se ha descrito,
la matriz colgena del primer cemento formado es el
resultado de las clulas asociadas al cemento y se ela-
bora antes de que se forme el ligamento periodontal;
por consiguiente, el colgeno es de origen local y, por
lo tanto, de derivacin intrnseca.
Cemento celular de fibras intrnsecas: tras haberse
formado, como mnimo, la mitad de la raz, los ce-
mentoblastos empiezan a formar un tipo menos mi-
neralizado de cemento que es caracterstico en cuanto
a que las fibrillas de colgeno que lo constituyen es-
tn producidas por los propios cementoblastos (fig. 5).
En todos los casos, el primer colgeno es depositado
sobre la superficie de dentina no mineralizada, de
forma que las fibrillas de ambas capas se entremez-
clan. Tal como ocurre en la formacin del cemento ace-
lular de fibras extrnsecas, los cementoblastos que for-
man el cemento celular de fibras intrnsecas tambin
elaboran un cierto nmero de protenas no colgenas
de la matriz que llenan los espacios entre las fibras de
colgeno, regulan el depsito de minerales e imparten
cohesin a la capa mineralizada. Una capa de matriz
no mineralizada, denominada cementoide, se esta-
blece en la superficie de la matriz de cemento mine-
ralizado, situndose el frente de mineralizacin en la
zona de unin entre las dos capas. A diferencia del os-
teoide, el cementoide no es tan regular ni tan fcil-
mente discernible. A medida que el proceso contina,
algunos cementoblastos se quedan atrapados en la ma-
triz que ellos mismos forman. Estas clulas atrapadas,
con una reducida actividad secretoria, se denominan
cementocitos y se asientan en lagunas. La organizacin
estructural de la matriz y la presencia de clulas en
sta otorgan al cemento celular de fibras intrnsecas
un aspecto seo. Durante la fase inicial las fibrillas de
colgeno son producidas rpidamente y depositadas
desordenadamente; sin embargo, la mayor parte de las
fibrillas se organiza a modo de haz, paralelamente a la
superficie radicular. Cuando el ligamento periodontal
queda organizado, puede formarse cemento alrededor
de algunos de los haces de fibras del ligamento perio-
dontal; de este modo, estos haces quedan incorpora-
dos en el cemento y se mineralizan parcialmente. En
los dientes humanos, la incorporacin de fibras del li-
gamento periodontal en el cemento celular de fibras
intrnsecas ocurre rara vez, esencialmente en el ce-
mento acelular de fibras extrnsecas que compone el
cemento celular estratificado mixto.
Cmo se sostiene el cemento sobre la dentina?
Es importante conocer el mecanismo de adheren-
cia del cemento a la dentina, no slo por un inters
biolgico, sino tambin por su posible aplicacin cl-
nica, ya que las alteraciones patolgicas y las inter-
venciones clnicas pueden influir sobre la naturaleza
de la superficie radicular expuesta y, por lo tanto, so-
bre la calidad de la nueva adherencia que se forma
cuando se deposita el cemento de reparacin. El me-
canismo por el cual se unen estos tejidos duros es
esencialmente el mismo para el cemento acelular de
fibras extrnsecas que para el cemento celular de fi-
bras intrnsecas. La mineralizacin del manto de den-
tina empieza internamente y no alcanza la superficie
hasta que las fibrillas de colgeno de la dentina y del
cemento hayan tenido tiempo de entremezclarse.
Luego se extiende por la superficie de la capa de den-
tina, a travs de la unin dentina-cemento y hacia el
cemento, dando como resultado una masa amalga-
mada de mineral. Mientras que la mineralizacin de
la dentina es iniciada por las vesculas de la matriz,
la posterior dispersin del depsito mineral est bajo
la influencia reguladora de las protenas no colge-
nas de la matriz. Desde una perspectiva biomecnica,
este ensamblaje parece ptimo para una fuerte unin
entre la dentina y el cemento. En el cemento acelu-
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
17
lar de fibras extrnsecas de los dientes de roedores, el
cemento se deposita sobre dentina mineralizada, por
lo cual la fusin de la dentina y el cemento resulta
imposible, y la unin entre estas capas es dbil. De
hecho, las secciones histolgicas de los dientes de ro-
edores a menudo muestran una separacin entre la
dentina y el cemento en el tercio cervical de la raz.
Si bien el procesado del tejido es considerado ha-
bitualmente como el responsable de la separacin ti-
sular en las secciones histolgicas, han surgido argu-
mentos que cuestionan esta interpretacin generali-
zada (16). Es interesante sealar que el cemento de
reparacin adhiere muy bien a la superficie de la raz
si una fase de resorcin precede al depsito de una
matriz nueva (14, 8), lo que indica que los odonto-
clastos preacondicionan favorablemente la superfi-
cie radicular. El preacondicionamiento qumico de la
superficie radicular con cidos o quelantes (fig. 7B)
es un paso que suele aplicarse en el tratamiento pe-
riodontal (43, 45). Despus de realizar varios proce-
dimientos regeneradores, frecuentemente se observa
una separacin tisular entre el cemento de repara-
cin y la superficie radicular tratada, lo que indica
una mala calidad de adherencia (8, 16) y la necesidad
de mejorar el acondicionamiento qumico de la su-
perficie radicular de los dientes afectados de perio-
dontitis.
Origen de los cementoblastos y de los fibroblastos
del ligamento periodontal
Existen todava algunos interrogantes que deben ser
resueltos, no slo para entender el proceso de la ce-
mentognesis, sino tambin y lo que es ms impor-
tante para disear planteamientos teraputicos dirigi-
dos a la prevencin y al tratamiento de las enfermedades
periodontales:
Cules son los precursores de los cementoblastos.
Si los cementoblastos son una poblacin celular dis-
tinta que expresa productos genticos nicos.
Si el cemento acelular y el cemento celular son te-
jidos diferentes.
Qu regula la formacin y el mantenimiento del li-
gamento periodontal frente al cemento, previniendo
as la fusin de la raz al hueso alveolar (anquilosis).
El punto de vista tradicional es que los precursores
de los cementoblastos y fibroblastos del ligamento pe-
riodontal residen en el folculo dental y que los facto-
res del entorno local regulan su capacidad para actuar
como cementoblastos que forman cemento radicular,
fibroblastos del ligamento periodontal u osteoblastos
que forman tejido seo (32). Se sostiene generalmente
que las clulas del folculo dental que se infiltran re-
Nanci y Bosshardt
18
Fig. 7. Microfotografa electrnica de transmisin de dien-
tes humanos afectados por periodontitis y tratados, de
forma conservadora, con raspado y alisado radicular, y de-
terminacin de la superficie radicular con cido etilendia-
minotetraactico (EDTA). A) Un epitelio de unin largo
(EUL) puede establecerse sobre la raz tratada. Ntese la
presencia de bacterias (cabezas de flecha) entre las clulas
epiteliales. B) el tratamiento con EDTA apunta a exponer
las fibrillas de colgeno (FC). AEFC: cemento acelular de fi-
bras extrnsecas.
EUL
FC
A
B
10 m 500 m
AEFC
AEFC
ciben una seal inductiva recproca de la dentina en
formacin y se diferencian en cementoblastos. Sin em-
bargo, cada vez se dispone de ms datos que apuntan
a que las clulas de la HERS pueden experimentar una
transformacin epitelial-mesenquimatosa en cemen-
toblastos durante el desarrollo (7). ste es un proceso
fundamental en biologa evolutiva, que se produce,
entre otros momentos, cuando las clulas ectodrmi-
cas migran fuera de la cresta neural y durante la fu-
sin del borde medial de los shelves palatinos. Diver-
sos hallazgos estructurales e inmunocitoqumicos
respaldan la hiptesis de que los cementoblastos de-
riven, al menos en parte, de clulas epiteliales trans-
formadas a partir de las clulas de la HERS. En mo-
delos de roedores, la formacin inicial del cemento
acelular tiene lugar en presencia de clulas epitelia-
les, y se ha mostrado que las clulas del rgano del
esmalte son capaces de generar productos mesen-
quimatosos tpicos, como el colgeno tipo I, la sialo-
protena sea y la osteopontina (10, 12, 13, 18, 48).
Se ha afirmado que el cemento acelular (primario)
y el cemento celular (secundario) estn producidos
por distintas poblaciones de clulas, cuyos compor-
tamientos espaciotemporales dan por resultado las di-
ferencias histolgicas caractersticas entre estos teji-
dos. No obstante, an hay controversias con respecto
a esta cuestin. Tambin se ha considerado la posibi-
lidad de que el cemento acelular de fibras extrnsecas
est formado por clulas derivadas de la HERS y que
el cemento celular de fibras intrnsecas est produ-
cido por clulas que derivan del folculo dental (68).
Los hallazgos experimentales respaldan el concepto
de que el ligamento periodontal es un depsito de las
clulas involucradas en la formacin del cemento, del
propio ligamento periodontal y del hueso alveolar (37);
sin embargo, no se ha determinado la naturaleza y la
localizacin precisa de las clulas progenitoras. Tam-
bin se desconoce si existen distintas lneas de clulas
precursoras para cada uno de los tres tejidos de soporte
o si los fibroblastos del ligamento periodontal, los ce-
mentoblastos y los osteoblastos surgen de un precur-
sor comn. La complejidad del ligamento periodontal
aumenta por el hecho de que contiene varios tipos de
clulas (subpoblaciones fibroblsticas, osteoblastos,
cementoblastos, clulas endoteliales, clulas perivas-
culares, clulas sanguneas y clulas epiteliales). Ade-
ms, hallazgos recientes tambin sugieren la presencia
de clulas con caractersticas de clulas madre (59). Una
revisin general de la bibliografa sobre el origen y la
diferenciacin celular ha sido recientemente efectuada
por Bosshardt (7). All se mencionan diversos hallazgos
que respaldan la hiptesis de que los cementoblastos
que producen tanto el cemento acelular de fibras ex-
trnsecas como el cemento celular de fibras intrnsecas
son fenotipos nicos, que difieren de los osteoblastos,
y propone un modelo que explica la diversificacin ce-
lular en el ligamento periodontal. Esta nueva teora pro-
pone que las clulas derivadas de la HERS desempe-
an un papel esencial en el desarrollo y el manteni-
miento del tejido, y que la regeneracin periodontal re-
capitula el desarrollo dentario. Las clulas que
descienden de la HERS pueden dar origen directamente
a clulas que formen nuevos tejidos de cemento y li-
gamento periodontal, o desempear un papel indirecto,
produciendo las molculas de sealizacin necesarias
para el reclutamiento y la diferenciacin de las clulas.
Es necesario profundizar los conocimientos acerca del
origen y la diferenciacin celular dentro del ligamento
periodontal, a fin de poder desarrollar terapias ms efi-
caces dirigidas al logro de una regeneracin periodon-
tal verdadera y significativa.
Los factores moleculares que regulan
la cementognesis
Protenas morfogenticas seas: Las protenas mor-
fogenticas seas (BMP) son miembros de la superfa-
milia del factor de transformacin del crecimiento
(TGF-) que actan a travs de los receptores serinci-
nasa y treonincinasa transmembrana. Estas molculas
de sealizacin tienen diversas funciones durante la
morfognesis y la diferenciacin celular y, en los dien-
tes, estn consideradas como parte del entramado de
las molculas de sealizacin epiteliales-mesenqui-
matosas que regulan el desarrollo de la corona. No es-
tn completamente esclarecidas las funciones de las
protenas morfogenticas seas en el desarrollo de la
raz, su supuestoa participacin en la sealizacin epi-
telial-mesenquimatosa, y las trayectorias de sealiza-
cin y los factores de transcripcin implicados en la
modulacin de su comportamiento. Sin embargo, se
sabe que varias de las protenas morfognicas seas,
como las BMP-2, BMP-4 y BMP-7, promueven la dife-
renciacin de los preosteoblastos y de las clulas pre-
cursoras putativas de los cementoblastos. En este con-
texto, las protenas morfgenas seas han sido
utilizadas con xito para inducir la regeneracin pe-
riodontal en cierto nmero de modelos experimenta-
les, aunque an no han sido utilizadas en la clnica.
Factores epiteliales: las mismas dos poblaciones de c-
lulas involucradas en la morfognesis de la corona, es
decir, el epitelio del esmalte y las clulas ectomesenqui-
matosas, tambin contribuyen a la formacin de la raz.
Por consiguiente, no sera sorprendente si algunas de las
mismas molculas de sealizacin implicadas en la mor-
fognesis de la corona tambin estuvieran activas du-
rante el desarrollo de la raz. Los posibles candidatos son
las protenas de la matriz del esmalte, la protena rela-
cionada con la hormona paratiroidea y los constituyen-
tes de la membrana basal. En el caso de las protenas de
la matriz del esmalte, el debate se centra en el hecho de
que no han sido sistemticamente detectadas a lo largo
de la raz en todas las especies y en todos los dientes.
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
19
Pese a ello, no se descarta su participacin en la forma-
cin de la raz. Algunas protenas podran, incluso, ser
secretadas transitoriamente en cantidades limitadas por
las clulas de la HERS en los estadios tempranos de la
formacin radicular, para influir sobre la diferenciacin
de odontoblastos y cementoblastos. Una expresin tan
limitada sera difcil de detectar.
Protenas de la matriz: segn ya se ha mencionado,
la sialoprotena sea y la osteopontina son constitu-
yentes fundamentales de la matriz del cemento, tanto
durante su desarrollo como durante su reparacin. Los
datos actualmente disponibles permiten suponer que
la osteopontina est involucrada en la regulacin del
crecimiento mineral, mientras que la sialoprotena
sea promueve la formacin mineral en la superficie
radicular (18, 49). Igualmente, pueden estar involu-
cradas en acontecimientos celulares mediante sus mo-
tivos de unin celular RGD. Dado que hasta la fecha
no se ha informado de ninguna anomala de desarro-
llo en ratones sin osteopontina, es probable que otras
protenas de la matriz no colgenas compensen la au-
sencia de osteopontina en estos animales.
La protena Gla sea (osteocalcina) es un marcador
de maduracin de los osteoblastos, odontoblastos
y cementoblastos que puede regular la extensin de
la mineralizacin. Hasta ahora no se ha comunicado
ningn problema de desarrollo y formacin en rato-
nes inactivados.
Factores de transcripcin: el Runx2 (factor de trans-
cripcin 2 relacionado al runt), tambin conocido
como Cbfa1 (factor 1 de unin al ncleo), y Osterix
(Osx), que es un objetivo posterior para el Cbfa1), han
sido identificados como los factores clave para la di-
ferenciacin de los osteoblastos. El Runx2 ha sido en-
contrado en clulas del folculo dental, clulas del li-
gamento periodontal, cementoblastos, cementocitos,
odontoblastos y ameloblastos. Basndose en las simi-
litudes propuestas con los osteoblastos, stos tambin
pueden estar involucradas en la diferenciacin de los
cementoblastos. Sin embargo, la expresin de Runx2
en clulas completamente diferenciadas sugiere fun-
ciones adicionales. Actualmente se estn investigando
los precisos factores que desencadenan la expresin
y/o la activacin de estos factores clave de transcrip-
cin; no obstante, las protenas genticas seas ya han
sido identificadas como factores que promueven la
expresin del Runx2.
Otros factores: entre otras molculas que pueden te-
ner una funcin reguladora en la diferenciacin y la
actividad de los cementoblastos se encuentran, den-
tro de los tejidos periodontales en desarrollo y ma-
duros, la fosfatasa alcalina, varios factores de creci-
miento (p. ej., el IGF, el TGF-y el factor de crecimiento
plaquetario), las metaloproteinasas y los proteoglu-
canos. Los ltimos son importantes para la formacin
de tejidos mineralizados, aunque no se ha establecido
una funcin especfica relacionada con la promocin
o la inhibicin de la diferenciacin de los cemento-
blastos. En la unin dentina-cemento se ha observado
una acumulacin de proteoglucanos, y se ha propuesto
que, juntamente con otras protenas no colgenas de
la matriz tales como la sialoprotena sea y la osteo-
pontina, pueden estar asociadas con la mineralizacin
inicial y la adherencia de las fibras (65).
Hace tiempo que se conoce la importancia de la fos-
fatasa alcalina para la formacin del cemento, particu-
larmente con respecto a las caractersticas distintivas en-
tre el cemento acelular de fibras extrnsecas y el cemento
celular de fibras intrnsecas. En los ratones sin el gen de
la fosfatasa alcalina inespecfica tisular o ratas tratadas
con biofosfonatos, la formacin de cemento acelular est
significativamente afectada mientras que el cemento ce-
lular parece desarrollarse con normalidad. Esto sugiere
diferencias en los tipos de clulas y/o los factores que
controlan el desarrollo de estas dos variedades de ce-
mento. En el ser humano, en casos de hipofosfatasia, ca-
racterizada por concentraciones muy bajas de fosfatasa
alcalina, la formacin de cemento parece ser escasa
o nula. En cambio, los ratones con mutaciones en los
genes que mantienen las concentraciones de pirofosfato
extracelular tales como ank y PC-1 y, en consecuen-
cia, con bajas concentraciones de pirofosfato, exhiben
mas cemento celular que las cras intactas de la misma
camada, incluso en los estadios tempranos del desarro-
llo de la raz (51). Estos hallazgos permiten suponer un
papel importante para la fosfatasa en el control de la ve-
locidad de formacin del cemento.
El ligamento periodontal
La mayor parte del ligamento periodontal es un te-
jido conectivo especializado, blando, situado entre el
cemento que cubre la raz del diente y el hueso que
forma la pared alveolar (ligamento alvolo-dental). Su
ancho oscila entre 0,15 y 0,38 mm, su porcin ms
delgada est situada alrededor del tercio medio de la
raz, y muestra una disminucin progresiva del gro-
sor con la edad. Es un tejido conectivo particularmente
bien adaptado a su funcin principal, sostener los
dientes en sus alvolos y, al mismo tiempo, permitir-
les resistir las considerables fuerzas de la masticacin.
Adems, el ligamento periodontal tiene la capacidad
de actuar como un receptor sensorial, necesario para
el apropiado posicionamiento de los maxilares du-
rante la masticacin y, lo que es an ms importante,
es un reservorio de clulas para la homeostasia tisu-
lar y para la reparacin o regeneracin.
Formacin del ligamento periodontal
El ligamento periodontal se forma dentro de la re-
gin del folculo dental, pero el momento exacto de
Nanci y Bosshardt
20
los acontecimientos asociados con el desarrollo de un
ligamento periodontal organizado vara entre espe-
cies, con cada familia de dientes y entre los dientes
primarios y los permanentes. Al principio, el espacio
del ligamento est ocupado por un tejido conectivo
desorganizado que se extiende entre el hueso y el ce-
mento. Este tejido es posteriormente remodelado, y la
matriz extracelular provisional se convierte en un sis-
tema de fibras organizadas como haces que se ex-
tienden entre las superficies del hueso y del cemento.
El tejido reorganizado puede ahora establecer conti-
nuidad a lo largo del espacio del ligamento y, de este
modo, asegurar la adhesin del diente al hueso. Pos-
teriormente, el movimiento causado por la erupcin
dentaria y el establecimiento de la oclusin modifi-
carn todava ms este sistema de adhesin inicial.
Constituyentes de las clulas del ligamento
periodontal y de la matriz extracelular
De modo similar al de otros tejidos conectivos, el li-
gamento periodontal consta de clulas y un comparti-
miento extracelular que comprende constituyentes co-
lgenos y no colgenos de la matriz. Entre las clulas se
encuentran osteoblastos y osteoclastos, fibroblastos,
restos de clulas epiteliales de Malassez, monocitos y
macrfagos, clulas mesenquimatosas indiferenciadas,
cementoblastos y odontoclastos. El compartimiento ex-
tracelular consiste principalmente en haces de fibras de
colgeno bien definidos incrustados en un material de
base amorfo, conocido como sustancia fundamental.
Fibroblastos: las principales clulas del ligamento pe-
riodontal son los fibroblastos. Aunque todos los fibro-
blastos se parecen microscpicamente, existen pobla-
ciones celulares heterogneas entre los diferentes tejidos
conectivos y tambin dentro del mismo tejido conec-
tivo. Los fibroblastos del ligamento periodontal estn ca-
racterizados por el rpido recambio del compartimiento
extracelular, en particular, el colgeno. Los fibroblastos
del ligamento periodontal son clulas grandes con un
extenso citoplasma que contiene una abundancia de or-
gnulos asociados con la sntesis y la secrecin de pro-
tenas. Tienen un citoesqueleto bien desarrollado y mues-
tran frecuentes adherencias y uniones intercelulares
comunicantes, lo que demuestra las exigencias funcio-
nales ejercidas sobre las clulas. Los fibroblastos del li-
gamento estn alineados a lo largo de la direccin ge-
neral de los haces de fibras y extienden los procesos
citoplsmicos que se producen a su alrededor. Las fibri-
llas de colgeno de los haces son continuamente remo-
deladas por los fibroblastos, que son capaces de sinteti-
zar y degradar el colgeno simultneamente.
Clulas epiteliales: las clulas epiteliales del ligamento
periodontal son residuos de las HERS de los restos epi-
teliales de Malassez. Se encuentran cerca del cemento,
como un conjunto de clulas que forman un entra-
mado epitelial, y parecen ser ms evidentes o abun-
dantes en las reas de furcacin. No se conoce exacta-
mente la funcin de estos residuos, pero se cree que
podran estar involucrados en la reparacin y regene-
racin periodontal (analizado en Bosshardt et al. [7]).
Clulas mesenquimatosas indiferenciadas: un impor-
tante constituyente celular del ligamento periodontal es
la clula mesenquimatosa indiferenciada, o clula pro-
genitora. El hecho de que se produzcan clulas nuevas
para el ligamento periodontal mientras que las clulas
del ligamento estn en un estado de equilibrio significa
que la eliminacin selectiva de las clulas por apoptosis
debe equilibrarse mediante la produccin de clulas nue-
vas. Durante la cicatrizacin de la lesin periodontal, el
ligamento periodontal contribuye con clulas no slo
para su propia reparacin, sino tambin para restaurar
el hueso y el cemento perdidos (5, 37). Recientemente
se han aislado del ligamento periodontal humano clu-
las con caractersticas de clulas madre (59).
Fibras: los colgenos predominantes del ligamento
periodontal son los colgenos de tipos I, III y XII, cu-
yas fibrillas tienen un dimetro medio relativamente
menor que el de las fibrillas de colgeno del tendn,
una diferencia que posiblemente refleje la semivida
relativamente corta del colgeno del ligamento y, por
lo tanto, el menor tiempo para un ensamblaje. La vasta
mayora de fibrillas de colgeno en el ligamento pe-
riodontal est organizada en haces de fibras bien de-
finidos, denominados fibras principales. Cada haz ase-
meja una cuerda empalmada; los filamentos pueden
ser continuamente remodelados mientras la fibra en
conjunto mantiene su arquitectura y su funcin. De
esta forma, los haces de fibras son capaces de adap-
tarse a las continuas sobrecargas a los que estn so-
metidos. Los extremos de los haces de fibras de col-
geno se insertan en el cemento o en el hueso. La porcin
insertada es denominada fibras de Sharpey. Las fibras
de Sharpey en el cemento acelular primario estn com-
pletamente mineralizadas; las que se encuentran en
el cemento celular y en el hueso estn, por lo gene-
ral, slo parcialmente mineralizadas en su periferia.
Otros haces de fibras (fibras del ligamento gingival)
se encuentran extendindose desde la regin cervical
de un diente hasta la regin cervical del diente adya-
cente (fibras del ligamento transeptal) y en la lmina
propia de la enca. stas, juntamente con las princi-
pales fibras del ligamento alvolo-dental, constituyen
el sistema de fibras del ligamento periodontal.
Fibras elsticas: existen tres tipos de fibras elsticas:
de elastina, de oxitaln y de elaunina. nicamente las
fibras de oxitaln estn presentes dentro del ligamento
periodontal; sin embargo, las fibras de elaunina tam-
bin pueden encontrarse asociadas a los haces de fi-
bras en el ligamento gingival.
Las fibras de oxitaln son haces de microfibrillas
que se dirigen ms o menos verticalmente desde la
superficie del cemento, formando una red bifurcada
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
21
tridimensional que rodea la raz y termina en el com-
plejo apical de arterias, venas y vasos linfticos. Tam-
bin estn asociadas con elementos neurales. Aunque
no se ha determinado completamente su funcin, se
piensa que regulan el flujo vascular en relacin con la
funcin del diente. Debido a que son elsticas, pue-
den expandirse en respuesta a las variaciones de la
presin, y estas variaciones se reflejan en las paredes
de las estructuras vasculares.
Protenas no colgenas de la matriz: varias prote-
nas no colgenas de la matriz producidas localmente
por las clulas residentes o aportadas por la circula-
cin se encuentran en el ligamento periodontal, en-
tre stas: la fosfatasa alcalina (31), los proteoglucanos
(33) y glucoprotenas como la undulina, la tenascina
y la fibronectina (69).
Sustancia fundamental: se estima que el 70 % de la
sustancia fundamental del ligamento periodontal es
agua, y se cree esta caracterstica tiene un efecto de-
terminante sobre la capacidad del diente para sopor-
tar las sobrecargas. Hay un aumento de fluidos tisu-
lares dentro de la matriz amorfa de la sustancia
fundamental en las reas de lesin e inflamacin.
Homeostasia y adaptacin del ligamento
periodontal a las exigencias funcionales
Una notable capacidad del ligamento periodontal es
que mantiene, relativamente, su anchura con el tiempo,
a pesar de que est presionado entre dos tejidos duros.
Existen pruebas convincentes que indican que las po-
blaciones de clulas dentro del ligamento periodontal,
tanto durante su desarrollo como durante su regenera-
cin, secretan molculas que pueden regular la exten-
sin de la mineralizacin y prevenir la fusin de la raz
dentaria con el hueso circundante (anquilosis). Entre
estas molculas, el equilibrio entre las actividades de la
sialoprotena sea y la osteopontina puede contribuir
a establecer y mantener una regin del ligamento pe-
riodontal no mineralizada. La protena Gla de la matriz
tambin est presente en los tejidos periodontales; de-
bido a su papel como inhibidor de la mineralizacin,
puede tambin actuar para preservar la anchura del li-
gamento periodontal. Al nivel celular, se ha documen-
tado que el Msx2 evita la diferenciacin ostegena de
los fibroblastos del ligamento periodontal, reprimiendo
la actividad de transcripcin de Runx2/Osf2 (67). Real-
mente, el Msx2 puede desempear una funcin central
evitando, por lo general, que los ligamentos y los ten-
dones se mineralicen (67). Tambin se ha sugerido que
los glucosaminoglucanos (39) o la protena de adhesin
al cemento RGD, una protena asociada al colgeno (52),
tambin pueden desempear una funcin en el man-
tenimiento del estado no mineralizado del ligamento
periodontal. En este punto, an no se ha esclarecido
cmo el ligamento periodontal, atrapado entre dos te-
jidos calcificados, permanece sin calcificarse, y ser ne-
cesario realizar nuevas investigaciones al respecto.
El ligamento periodontal tambin tiene la capacidad
de adaptarse a los cambios funcionales. Cuando la de-
manda funcional aumenta, la anchura del ligamento pe-
riodontal puede aumentar hasta un 50 %, y los haces de
fibras tambin pueden aumentar notablemente en gro-
sor. En cambio, una reduccin en la funcin conduce al
estrechamiento del ligamento y a una disminucin del
nmero y el grosor de los haces de fibras. Estas modifi-
caciones funcionales del ligamento periodontal tambin
ocasionan los correspondientes cambios adaptativos en
el cemento y el hueso alveolar circundantes.
El hueso alveolar
El proceso alveolar es aquel hueso de los maxilares
que contiene las cavidades (alvolos) para los dientes.
Consta de placas corticales externas (vestibular, lin-
gual y palatina) de hueso compacto, una esponjosa
central y el hueso que reviste los alvolos (hueso al-
veolar). La placa cortical y el hueso que reviste el al-
volo entran en contacto a la altura de la cresta alve-
olar. El hueso que reviste el alvolo es denominado
hueso del haz, porque proporciona adhesin a los ha-
ces de fibras del ligamento periodontal.
Las placas corticales constan de capas de superficie
(laminillas) de hueso de fibra fina mantenidas por sis-
temas haversianos. Generalmente son ms delgadas en
el maxilar superior y ms gruesas en la cara vestibular
de los premolares y molares mandibulares. El hueso tra-
becular (o esponjoso) que ocupa la parte central del pro-
ceso alveolar tambin consta de hueso dispuesto en l-
minas, con sistemas haversianos presentes en las
trabculas ms grandes. La mdula amarilla, rica en c-
lulas adiposas, generalmente rellena los espacios inter-
trabeculares, aunque algunas veces tambin puede ha-
ber un poco de mdula roja o hematopoytica. En la
regin de los dientes anteriores no hay hueso trabecu-
lar y, en este caso, la placa cortical y el hueso alveolar
estn fusionados. La parte central de este complejo, en
cuanto al sostn dentario, es el hueso del haz, que consta
de capas sucesivas de haces de fibra intrnseca, con un
recorrido ms o menos paralelo al alvolo. Incrustados
dentro de este hueso del haz, casi perpendiculares a su
superficie, estn los extremos (fibras de Sharpey) de los
haces de fibras extrnsecas de colgeno del ligamento
periodontal (que, al igual que en el cemento celular de
fibras intrnsecas y en el cemento celular estratificado
mixto, estn mineralizados slo en su periferia). Debido
a que el diente est constantemente sometido a movi-
mientos menores y a que el hueso alveolar debe res-
ponder a la demanda funcional ejercida sobre l por las
fuerzas de la masticacin, el hueso de la pared del al-
volo se remodela constantemente y su organizacin
estructural vara a lo largo de la pared (56). La presen-
Nanci y Bosshardt
22
cia de un hueso alveolar que recorre todo el alvolo den-
tario separa anatmica y funcionalmente el hueso de
soporte y el ligamento periodontal. La organizacin del
proceso alveolar es otro ejemplo ms de la relacin que
existe entre la estructura y la funcin en el periodoncio.
Mientras que los procesos de formacin y regula-
cin del hueso alveolar son similares a los que se ob-
servan en otras zonas anatmicas, el hueso alveolar
tiene caractersticas distintivas, porque su recambio
celular es muy rpido y se pierde en ausencia de un
diente. Estas dos caractersticas sugieren que los me-
canismos reguladores locales son particularmente im-
portantes en el caso del hueso alveolar. Tambin de-
muestran claramente la interdependencia de los
tejidos periodontales y subrayan el importante hecho
de que los tejidos periodontales funcionan como una
unidad.
El proceso de remodelado del hueso alveolar es esen-
cialmente similar al del hueso en general (56). Sin em-
bargo, la resorcin es asincrnica, de modo que la in-
sercin del ligamento periodontal se pierde solamente
de forma focal y por cortos perodos de tiempo. Du-
rante la migracin dentaria, la distribucin de la fuerza
es tal que el hueso perdido a causa de la resorcin en
una superficie del alvolo dentario es equilibrado por
la formacin de hueso a lo largo de la superficie
opuesta. Este equilibrio seo, juntamente con el con-
tinuo depsito de cemento durante toda la vida, ac-
tan para mantener una relacin ms o menos cons-
tante entre la superficie de la raz y la de la cavidad
alveolar. Todava no se conocen exactamente todos los
factores que desencadenan los diversos aconteci-
mientos en la homeostasia periodontal. La compren-
sin de estos procesos sera de gran utilidad, pues per-
mitira sacar provecho de los acontecimientos que se
producen durante el tratamiento ortodncico, que
esencialmente representa una circunstancia donde los
lmites de la fisiologa normal se distienden.
Tal como se ha mencionado, en el ligamento pe-
riodontal hay clulas progenitoras que pueden dife-
renciarse en osteoblastos para el mantenimiento fi-
siolgico del hueso alveolar y, probablemente,
tambin para su reparacin. Dado que los datos dis-
ponibles permiten suponer, en el ligamento perio-
dontal, un precursor para los cementoblastos, los fi-
broblastos del ligamento periodontal y las clulas
seas (7), sera muy beneficioso identificar las sea-
les que guan la diferenciacin celular en cada una
de estas trayectorias, a fin de disear terapias rege-
neradoras dirigidas.
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
23
Fig. 8. Microfotografas electrnicas ptica (A) y de trans-
misin (B) de dientes humanos afectados por periodonti-
tis. A) La bolsa periodontal (BP) ocupa el espacio entre la
placa bacteriana (PB) adherida a la superficie radicular y
el epitelio (EP) que reviste el espacio de la bolsa. Ntese la
gran cantidad de vasos sanguneos (VS) en el epitelio de la
bolsa. B) Unos das despus del raspado y alisado radicu-
lar los agentes patgenos periodontales pueden reestable-
cer una biopelcula bacteriana sobre la superficie de den-
tina (D) expuesta.
PB
PB
BP
D
VS
EP
10 m
A
B
1 m
Alteraciones patolgicas
de la estructura y la funcin
de los tejidos periodontales
La gingivitis y la periodontitis (figs. 7 y 8) son en-
fermedades infecciosas que afligen a un alto porcen-
taje de la poblacin, incluso a individuos jvenes. En
su documento FY2003 Fact Sheet, la American Asso-
ciation for Dental Research inform que el 48 % de
los adultos de edad comprendida entre los 35 y 44
aos presenta inflamacin de la enca (gingivitis), y el
22 % sufre una enfermedad periodontal destructiva,
una importante causa de perdida dentaria. Adems,
se ha comprobado que las enfermedades periodonta-
les crnicas estn relacionadas con las principales en-
fermedades sistmicas, tales como las enfermedades
cardiovasculares y pulmonares (4, 23, 28, 53). Aunque
las bacterias son imprescindibles para que se desa-
rrolle la periodontitis, el hecho de que se desarrolle
en grados variables en individuos diferentes sugiere
una etiologa multifactorial. Todas las manifestacio-
nes de periodontitis, no obstante, parecen tener una
serie de acontecimientos subyacentes comunes que
conducen a la destruccin del tejido y a la prdida de
la insercin dentaria.
El epitelio de unin, en virtud de su singularidad
estructural y funcional, proporciona una barrera muy
eficaz contra los agentes patgenos periodontales
y sus productos metablicos. Sin embargo, dichos
agentes, en especial Porphyromonas gingivalis, pue-
den perturbar su integridad, al permiir la disemina-
cin subgingival de bacterias y sus antgenos (9, 19,
35, 55). La consecuente respuesta inflamatoria con-
duce a la degradacin del tejido conectivo subyacente,
primero alrededor de los vasos sanguneos y luego en
zonas adyacentes, ocasionando la desintegracin es-
tructural y funcional de la enca.
Unos de los primeros cambios de la periodontitis
son la migracin del epitelio de unin a lo largo de la
superficie radicular y su elongacin, lo que conduce
a la formacin de un epitelio de unin largo y de una
bolsa gingival. Esta alteracin estructural va acompa-
ada de un cierto nmero de cambios funcionales. La
direccin de la migracin de los neutrfilos y del flujo
del exudado crevicular a travs del epitelio cambia
Nanci y Bosshardt
24
Fig. 9. Ilustracin esquemtica del sistema RANK-RANKL-osteoprotegerina
Progenitor
de osteoclastos
Linfocito T
Proliferacin
Diferenciacin
RANK
sOPG
sRANKL
mOPG
M-CSF
Progenitor
meieloide
Progenitor
hematopoytico
Osteoblasto
Clula del estroma
Linfocito B
Preosteoclasto
Osteoclasto
activo
Estimuladores:
TGF-
17--estradiol
Estimuladores:
PTH
1,25(OH
2
)D
3
PGE
2
Il-1, 6, 11, 17
TNF
IFN-
IFNR
mRANKL
Fusin de
supervivencia
Activacin
drsticamente, ya que la superficie libre del epitelio
es ahora desplazada desde el fondo del surco hasta la
superficie de la raz. Asimismo, la superficie libre au-
menta en tamao y, por lo tanto, est expuesta a ms
placa bacteriana.
La naturaleza del tejido conectivo con el que el epi-
telio de unin est en contacto influye, segn se cree,
sobre su desarrollo. Esta lnea de razonamiento puede
servir para explicar, de modo similar, la formacin de
un epitelio de unin largo. El epitelio de unin nece-
sita, para establecerse, un cierto entorno de tejido
conectivo cuyas caractersticas especficas no han
sido todava definidas que, en un periodoncio sano,
habitualmente se encuentra cerca de la porcin cer-
vical del diente. Cuando se instaura la gingivitis, el te-
jido conectivo que bordea el epitelio de unin es con-
tinuamente alterado por la respuesta inflamatoria. Por
consiguiente, necesita migrar a mayor profundidad
a lo largo de la superficie de la raz para encontrar una
estructura de tejido conectivo que est suficiente in-
tacta y que sea capaz de enviar seales al epitelio para
detener su movimiento en direccin hacia abajo, for-
mar una insercin epitelial funcional y adherir a la su-
perficie dentaria.
Las bacterias causan la destruccin tisular indirec-
tamente, mediante la exacerbacin de la respuesta in-
munitaria del hospedador. Teng (62) y Yamazaki et al.
(66) han revisado los recientes avances en las respues-
tas inmunitarias adquiridas que en el contexto de pro-
gresin de la enfermedad periodontal involucran a los
linfocitos B, los linfocitos T y mediadores inflamatorios.
Se sabe que diversas citocinas proinflamatorias y fac-
tores de crecimiento, especialmente la IL-1 y el factor
de necrosis tumoral (TNF-), estn asociados con la
resorcin del hueso (fig. 9). El remodelado seo nor-
mal depende de un delicado equilibrio entre la forma-
cin de hueso y su resorcin. El receptor activador del
factor nuclear B (RANK) y su ligando el RANKL, miem-
bros de la familia de receptores del factor de necrosis
tumoral, estn directamente implicados en la diferen-
ciacin de los precursores de los osteoclastos y en la
activacin y supervivencia de los osteoclastos (fig. 9).
El RANKL es expresado por las clulas del estroma de
la mdula sea, los osteoblastos y los fibroblastos, mien-
tras que el RANK es expresado por los precursores de
los osteoclastos y por osteoclastos maduros. La unin
del RANK y del RANKL induce la diferenciacin y la ac-
tividad de los osteoclastos. La osteoprotegerina, que
est producida por las clulas del estroma medular, por
los osteoblastos y por los fibroblastos del ligamento pe-
riodontal, compite por esta unin y funciona como un
receptor seuelo soluble para el RANKL. Por consi-
guiente, la osteoprotegerina es un inhibidor natural de
la diferenciacin y la activacin de los osteoclastos.
Cualquier interferencia con este sistema puede cam-
biar de direccin este equilibrio hacia un aumento de
la formacin o la resorcin de hueso. Se ha demostrado
que las citocinas proinflamatorias como la IL-1 y el
TNF-, dos protagonistas muy importantes en la pr-
dida de hueso periodontal, regulan la expresin de
RANKL y osteoprotegerina. Adems, los linfocitos T
tambin expresan RANKL, que unindose directamente
al RANK en la membrana celular de los progenitores
de osteoclastos, preosteoclastos y osteoclastos, estimula
tanto la diferenciacin celular como la activacin de
las clulas del linaje de los osteoclastos. Por lo tanto, el
giro de la homeostasia sea hacia la resorcin sea en
la periodontitis puede ser ocasionado por las citocinas
proinflamatorias que regulan la expresin de RANKL
tanto en las clulas mesenquimatosas como en espe-
cficos linfocitos T activados. Al respecto, se ha demos-
trado que existe una disminuida prdida de hueso al-
veolar tras la infeccin bucal con P. gingivalis en ratones
que carecen de linfocitos T (2). El descubrimiento de
este sistema regulador que conecta la biologa sea con
la biologa celular inmunitaria (29, 30, 34) ha abierto
nuevas posibilidades teraputicas, como la inhibicin
de la interaccin entre el RANK y el RANKL mediante
la aplicacin local de osteoprotegerina.
Debido a la velocidad excepcionalmente alta de re-
cambio de colgeno en el ligamento periodontal, cual-
quier interferencia con la funcin de los fibroblastos
por enfermedad produce rpidamente una prdida
del tejido de soporte del diente. Es interesante que, en
las situaciones inflamatorias, como aquellas asocia-
das con las enfermedades periodontales, haya un au-
mento de la expresin de metaloproteinasas de la ma-
triz que destruyen agresivamente el colgeno (54). Por
lo tanto, las terapias para controlar la destruccin ti-
sular pueden incluir moduladores del hospedador que
tengan la capacidad de inhibir las metaloproteinasas
de la matriz.
Cuando la inflamacin alcanza la superficie de la
raz, puede producirse resorcin, la que da por resul-
tado la excavacin de la raz. Es posible que este pro-
ceso destructivo implique cierto desequilibrio en el
sistema RANK-RANKL-osteoprotegerina (fig. 9) (7). La
resorcin radicular se produce de forma bastante ha-
bitual en la gingivitis hiperplsica y, con menor fre-
cuencia, adyacente a lesiones inflamatorias en el li-
gamento periodontal. Las fracturas y microfracturas
en la porcin superficial de la raz pueden facilitar la
invasin de las bacterias o la difusin de los produc-
tos bacterianos hacia la raz. El cemento y la dentina
daados tambin pueden actuar como reservorios
bacterianos, a partir de los cuales se puede producir
la recolonizacin de las superficies radiculares raspa-
das y alisadas. Como aspecto positivo, la exposicin
al entorno bucal a menudo conduce al establecimiento
de una zona hipermineralizada en la capa de cemento
superficial. Los cristales minerales en esta capa su-
perficial son muy resistentes a la desmineralizacin
con cido, lo que lentifica el avance de las lesiones de
caries.
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
25
Coclusiones
En el prefacio de Periodontology 2000 (57), el editor
Schroeder afirma lo siguiente: Es necesario com-
prender la arquitectura de los tejidos (humanos), a fin
de que las mentes creativas puedan formular los inte-
rrogantes biolgicos pertinentes. Realmente, es fun-
damental conocer cmo se desarrolla la estructura ti-
sular y la relacin estructura-funcin, para entender el
proceso patolgico y disear estrategias teraputicas
eficaces, especialmente cuando se produce destruc-
cin tisular y, en consecuencia, una prdida funcional.
El presente captulo ha sido organizado teniendo pre-
sente este objetivo, y con la esperanza de proporcio-
nar a los investigadores una base slida para el diseo
de terapias regeneradoras. Un ejemplo especialmente
bueno de reparacin tisular inducida que puede no
dar resultados funcionales ptimos es la estimulacin
de la formacin de cemento celular de fibras intrnse-
cas para la reparacin radicular. Como se ha analizado
antes, el depsito de cemento celular de fibras intrn-
secas en las races afectadas de periodontitis tratadas
de diversas maneras no es la mejor manera de lograr
la insercin, pues cuando el cemento se deposita en-
cima de una superficie mineralizada no interacta con
la matriz calcificada preexistente y, por lo tanto, est
sujeto a un posible desprendimiento. Este desprendi-
miento, sin embargo, tambin puede deberse a pro-
piedades inadecuadas de la superficie de la raz.
La primera lnea de defensa es el epitelio de unin.
Su alteracin estructural es, claramente, el primer paso
hacia la progresin de la enfermedad. Se ha prestado
relativamente poca atencin a los factores que de-
sencadenan su formacin y a la composicin de la es-
tructura tipo lmina basal que media su adherencia
a la superficie de la raz. Podran utilizarse los cons-
tituyentes de esta capa adhesiva para lentificar el cre-
cimiento hacia abajo del epitelio de unin y su sepa-
racin de la superficie dentaria? Una vez que la
enfermedad avanza ms all de la zona epitelial y se
disemina a los elementos del tejido conectivo del pe-
riodoncio, la regeneracin se complica por el hecho
de que ahora estn implicados tres tejidos: el cemento,
el ligamento periodontal y el hueso. Aunque, en prin-
cipio, cualquiera de ellos puede ser reconstruido, es
preciso recordar que la funcin de la insercin den-
taria requiere que la estructura de los tres tejidos sea
restaurada de modo que se mantenga un adecuado
equilibrio entre ellos. En el caso del cemento, adems
de la cantidad, el tipo de cemento tambin ser un
elemento clave.
Todava se desconoce si el precursor para las clu-
las periodontales es ectodrmico o ectomesenquima-
toso, o si hay ms de un precursor. No obstante, esto
puede ser una cuestin de semntica, ya que las c-
lulas ectomesenquimatosas tambin son de origen ec-
todrmico. El hecho de que las terapias periodonta-
les actuales estimulen principalmente la formacin de
cemento celular de fibras intrnsecas y que el cemento
acelular de fibras extrnsecas y el cemento celular de
fibras intrnsecas puedan verse particularmente afec-
tados por las enfermedades o condiciones experi-
mentales, ciertamente sugiere que pueden ser provo-
cadas trayectorias celulares especficas y que existen
diferentes vas de sealizacin. Es importante reco-
nocer este hecho, a fin de lograr una regeneracin pe-
riodontal eficaz y especficamente dirigida.
En estos ltimos aos, se ha prestado mucha aten-
cin a las protenas de la matriz del esmalte, y los en-
sayos para su utilizacin en reparacin periodontal
han mostrado resultados clnicos prometedores. No
obstante, debe an investigarse la variabilidad en los
resultados clnicos y los posibles beneficios con res-
pecto al desbridamiento con colgajo abierto o la re-
generacin tisular guiada. Pueden obtenerse resulta-
dos clnicos relativamente similares con enfoques
teraputicos muy diferentes, lo que indica que estas
protenas de la matriz del esmalte pueden funcionar
indirectamente. Ciertamente, todava no se dispone
de ninguna prueba de que realmente sean los prin-
cipales protagonistas en la formacin de la raz. Desde
luego, todava no se ha comunicado ningn defec-
to radicular importante en ratones sin amelogenina
y ameloblastina y en ratones transgnicos. Esto no sig-
nifica que no deban usarse protenas de la matriz del
esmalte para el tratamiento periodontal, sino que si
se comprendiera mejor el mecanismo por el cual ejer-
cen su influencia, se podran utilizar estas u otras pro-
tenas de un modo ms eficaz.
Periodontology 2000, Vol. 40, 2006, 11-28
Bibliografa
1. Baba O, Qin C, Brunn JC, Jones JE, Wygant JN, Mcintyre
BW, Butler WT. Detection of dentin sialoprotein in rat
periodontium. Eur J Oral Sci 2004: 112: 163170.
2. Baker PJ, Howe L, Garneau J, Roopenian DC. T cell
knockout mice have diminished alveolar bone loss after
oral infection with Porphyromonas gingivalis. FEMS
Immunol Med Microbiol 2002: 34: 4550.
3. Bartold PM, Walsh LJ, Narayanan S. Molecular cell biology
of the gingiva. Periodontol 2000 2000: 24: 2855.
4. Beck JD, Slade G, Offenbacher S. Oral disease, cardiovas-
cular disease and systemic inflammation. Periodontol 2000
2000: 23: 110120.
5. Beertsen W, Van Den Bos T, Everts V. Continuous growth of
acellular extrinsic fiber cementum: a review. Acta Med Dent
Helv 1997: 2: 103115.
6. Berkovitz BK. Periodontal ligament: structural and clinical
correlates. Dent Update 2004: 31: 4650.
7. Bosshardt DD. Are cementoblasts a subpopulation of
osteoblasts or a unique phenotype? J Dent Res 2005: 84:
390406.
8. Bosshardt DD, Degen T, Lang NP. Sequence of protein
expression of bone sialoprotein and osteopontin at the
Nanci y Bosshardt
26
developing interface between repair cementum and dentin
in human deciduous teeth. Cell Tissue Res 2005: 320:
399407.
9. Bosshardt DD, Lang NP. The junctional epithelium: from
health to disease. J Dent Res 2005: 84: 920.
10. Bosshardt DD, Nanci A. Immunodetection of enamel-,
cementum-related (bone) proteins at the enamel-free area
and cervical portion of the tooth in rat molars. J Bone Miner
Res 1997: 12: 367379.
11. Bosshardt DD, Nanci A. Immunolocalization of epithelial
and mesenchymal matrix constituents in association with
inner enamel epithelial cells. J Histochem Cytochem 1998:
46: 135142.
12. Bosshardt DD, Nanci A. Immunocharacterization of
cementicles and ectopic enamel. J Dent Res (Spec Iss) 2000:
79: 563.
13. Bosshardt DD, Nanci A. Hertwigs epithelial root sheath
enamel matrix proteins and initiation of cementogenesis in
porcine teeth. J Clin Periodontol 2004: 31: 184192.
14. Bosshardt DD, Schroeder HE. How repair cementum
becomes attached to the resorbed roots of human perma-
nent teeth. Acta Anat (Basel) 1994: 150: 253266.
15. Bosshardt DD, Schroeder HE. Cementogenesis reviewed: a
comparison between human premolars and rodent molars.
Anat Rec 1996: 245: 267292.
16. Bosshardt DD, Sculean A, Windisch P, Pjetursson BE, Lang
NP. Effects of enamel matrix proteins on tissue formation
along the roots of human teeth. J Periodontal Res 2005: 40:
158167.
17. Bosshardt DD, Selvig KA. Dental cementum: the dynamic
tissue covering of the root. Periodontol 2000 1997: 13: 4175.
18. Bosshardt DD, Zalzal S, Mckee MD, Nanci A. Develop-
mental appearance, distribution of bone sialoprotein,
osteopontin in human and rat cementum. Anat Rec 1998:
250: 121.
19. Brien-Simpson NM, Veith PD, Dashper SG, Reynolds EC.
Porphyromonas gingivalis gingipains: the molecular teeth
of a microbial vampire. Curr Protein Pept Sci 2003: 4:
409426.
20. Butler WT, Brunn JC, Chunlin C, Mckee MD. Cell differ-
entiation (cost-wg3); extracellular matrix proteins, the
dynamics of dentin formation. Connect Tissue Res 2002: 43:
301307.
21. Bye FI, Caffesse RG, Nasjleti CE. The effect of different
plaque control modalities on the keratinizing potential of
the sulcular epithelium in monkeys. J Periodontol 1980: 51:
632641.
22. Caffesse RG, Kornman KS, Nasjleti CE. The effect of
intensive antibacterial therapy on the sulcular environ-
ment in monkeys. Part II. Inflammation, mitotic activity
and keratinization of the sulcular epithelium. J Periodontol
1980: 51: 155161.
23. Dave S, Batista ELJ, Van Dyke TE. Cardiovascular disease
and periodontal diseases: commonality and causation.
Compend Contin Educ Dent 2005: 25: 2637.
24. Feng JQ, Huang H, Lu Y, Ye L, Xie Y, Tsutsui TW, Kunieda T,
Castranio T, Scott G, Bonewald LB, Mishina Y. The dentin
matrix protein 1 (dmp1) is specifically expressed in min-
eralized, butnotsoft tissues during development. J Dent Res
2003: 82: 776780.
25. Fong CD, Hammarstrom L. Expression of amelin and
amelogenin in epithelial root sheath remnants of fully
formed rat molars. Oral Surg Oral Med Oral Pathol Oral
Radiol Endod 2000: 90: 218223.
26. Fong CD, Hammarstrm L, Lundmark C, Wurtz T, Slaby I.
Expression patterns of RNAs for amelin and amelogenin in
developing rat molars and incisors. Adv Dent Res 1996: 10:
195200.
27. Frank DE, Carter WG. Laminin 5 deposition regulates ker-
atinocyte polarization and persistent migration. J Cell Sci
2004: 117: 13511363.
28. Garcia RI, Henshaw MM, Krall EA. Relationship between
periodontal disease and systemic health. Periodontol 2000
2001: 25: 2136.
29. Goldring SR. Inflammatory mediators as essential elements
in bone remodeling. Calcif Tissue Int 2003: 73: 97100.
30. Grcevic D, Katavic V, Lukic IK, Kovacic N, Lorenzo JA,
Marusic A. Cellular and molecular interactions between
immune system and bone. Croat Med J 2001: 42: 384392.
31. Groeneveld MC, Van Den Bos T, Everts V, Beertsen W. Cell-
bound and extracellular matrix-associated alkaline phos-
phatase activity in rat periodontal ligament. J Periodontal
Res 1996: 31: 7379.
32. Grzesik WJ, Narayanan AS. Cementum and periodontal
wound healing and regeneration. Crit Rev Oral Biol Med
2002: 13: 474484.
33. Hakkinen L, Oksala O, Salo T, Rahemtulla F, Larjava H.
Immunohistochemical localization of proteoglycans in
human periodontium. J Histochem Cytochem 1993: 41:
16891699.
34. Hofbauer LC, Heufelder AE. Role of receptor activator of
nuclear factor-jB ligand and osteoprotegerin in bone cell
biology. J Mol Med 2001: 79: 243253.
35. Kadowaki T, Yamamoto K. Suppression of virulence of
Porphyromonas gingivalis by potent inhibitors specific for
gingipains. Curr Protein Pept Sci 2003: 4: 451458.
36. Karring T, Lang NP, Loe H. The role of gingival connective
tissue in determining epithelial differentiation. J Perio-
dontal Res 1975: 10: 111.
37. Karring T, Nyman S, Gottlow J, Laurell L. Development
of the biological concept of guided tissue regeneration
animal and human studies. Periodontol 2000 1993: 1: 2635.
38. Karring T, Ostergaard E, Loe H. Conservation of tissue
specificity after heterotopic transplantation of gingiva and
alveolar mucosa. J Periodontal Res 1971: 6: 282293.
39. Kirkham J, Brookes SJ, Shore RC, Bonass WA, Robinson C.
The effect of glycosylaminoglycans on the mineralization
of sheep periodontal ligament in vitro. Connect Tissue Res
1995: 33: 2329.
40. Lindskog S. Formation of intermediate cementum. I. Early
mineralization of aprismatic enamel and intermediate
cementum in monkey. J Craniofac Genet Dev Biol 1982: 2:
147160.
41. Lindskog S. Formation of intermediate cementum. II. A
scanning electron microscopic study of the epithelial rooth
sheath of Hertwig in monkey. J Craniofac Genet Dev Biol
1982: 2: 161169.
42. Lindskog S, Hammarstrm L. Formation of intermediate
cementum. III:
3
H-tryptophan and
3
H-proline uptake into
the epithelial root sheath of Hertwig in vitro. J Craniofac
Genet Dev Biol 1982: 2: 171177.
43. Lowenguth RA, Blieden TM. Periodontal regeneration: root
surface demineralization. Periodontol 2000 1993: 1: 5468.
44. Macdougall M, Gu TT, Luan XG, Simmons D, Chen JK.
Identification of a novel isoform of mouse dentin matrix
protein. 1. Spatial expression in mineralized tissues. J Bone
Miner Res 1998: 13: 422431.
45. Mariotti A. Efficacy of chemical root surface modifiers in
the treatment of periodontal disease: a systematic review.
Ann Periodontol 2003: 8: 205226.
Estructuras de los tejidos periodontales en el individuo sano y en el individuo enfermo
27
46. Marks SCJR, Mckee MD, Zalzal S, Nanci A. The epithelial
attachment and the dental junctional epithelium: ultra-
structural features in porcine molars. Anat Rec 1994: 238:
114.
47. Matias MA, Li H, Young WG, Bartold PM. Immunohisto-
chemical localisation of extracellular matrix proteins in the
periodontium during cementogenesis in the rat molar.
Arch Oral Biol 2003: 48: 709716.
48. Mizuno N, Shiba H, Mouri Y, Xu W, Kudoh S, Kawaguchi H,
Kurihara H. Characterization of epithelial cells derived
from periodontal ligament by gene expression patterns of
bone-related and enamel proteins. Cell Biol Int 2005:
29:111117.
49. Nanci A. Content and distribution of noncollagenous
matrix proteins in bone and cementum: relationship to
speed of formation and collagen packing density. J Struct
Biol 1999: 126: 256269.
50. Nanci A, Somerman M. The periodontium. In: Nanci A, ed.
Ten Cates Oral histology: development, structure, and
function. St. Louis: Harcourt Health Sciences, 2003.
51. Nociti FH, Berry JE, Foster BI, Gurley KA, Kingsley DM,
Takata T, Miyauchi M, Somerman MJ. Cementum: a
phosphate-sensitive tissue. J Dent Res 2002: 81: 817821.
52. Ohno S, Doi T, Fujimoto K, Ijuin C, Tanaka N, Tanimoto K,
Honda K, Nakahara M, Kato Y, Tanne K. Rgd-Cap (Beta Ig-
H3) exerts a negative regulatory function on mineralization
in the human periodontal ligament. J Dent Res 2002: 81:
822825.
53. Page RC, Offenbacher S, Schroeder HE, Seymour GJ,
Kornman KS. Advances in the pathogenesis of periodonti-
tis: summary of developments, clinical implications and
future directions. Periodontol 2000 1997: 14: 216248.
54. Potempa J, Banbula A, Travis J. Role of bacterial protein-
ases in matrix destruction and modulation of host
responses. Periodontol 2000 2000: 24: 153192.
55. Potempa J, Travis J. Porphyromonas gingivalis proteinases
in periodontitis: a review. Acta Biochim Pol 1996: 43: 455465.
56. Saffar JI, Lasfargues JJ, Cherruau M. Alveolar bone and the
alveolar process: the socket that is never stable. Periodontol
2000 1997: 13: 7690.
57. Schroeder HE. Biological structure of the normal and dis-
eased periodontium preface. Periodontol 2000 1997: 13:
77.
58. Schroeder HE, Listgarten MA. The gingival tissues: the
architecture of periodontal protection. Periodontol 2000
1997: 13: 91120.
59. Seo BM, Miura M, Gronthos S, Bartold PM, Batouli S,
Brahim J, Young M, Robey PG, Wang CY, Shi S. Investiga-
tion of multipotent postnatal stem cells from human per-
iodontal ligament. Lancet 2004: 364: 149155.
60. Slavkin HC, Bessem C, Fincham AG, Bringas PJR, Santos V,
Snead MI, Zeichner-David M. Human and mouse cemen-
tum proteins immunologically related to enamel proteins.
Biochim Biophys Acta 1989: 991: 1218.
61. Sodek J, Ganss B, Mckee MD. Osteopontin. Crit Rev Oral
Biol Med 2000: 11: 279303.
62. Teng YT. The role of acquired immunity and periodontal
disease progression. Crit Rev Oral Biol Med2003: 14:237252.
63. Thomas CH, Mcfarland CD, Jenkins MI, Rezania A, Steele
JG, Healy KE. The role of vitronectin in the attachment and
spatial distribution of bone-derived cells on materials with
patterned surface chemistry. J Biomed Mater Res 1997: 37:
8193.
64. Wu DY, Ikezawa K, Parker T, Saito M, Narayanan AS.
Characterization of a collagenous cementum-derived
attachment protein. J Bone Miner Res 1996: 11: 686692.
65. Yamamoto T, Domon T, Takahashi S, Arambawatta AKS,
Wakita M. Immunolocalization of proteoglycans and bone-
related noncollagenous glycoproteins in developing acel-
lular cementum of rat molars. Cell Tissue Res 2004: 317:
299312.
66. Yamazaki K, Yoshie H, Seymour GJ. T cell regulation of the
immune response to infection in periodontal diseases.
Histol Histopathol 2003: 18: 889896.
67. Yoshizawa T, Takizawa F, Iizawa F, Ishibashi O, Kawashima
H, Matsuda A, Endo N, Kawashima H. Homeobox protein
Msx2 acts as a molecular defense mechanism for pre-
venting ossification in ligament fibroblasts. Mol Cell Biol
2004: 24: 34603472.
68. Zeichner-David M, Oishi K, Su Z, Zakartchenko V, Chen LS,
Arzate H, Bringas P Jr. Role of Hertwigs epithelial root
sheath cells in tooth root development. Dev Dyn 2003: 228:
651663.
69. Zhang X, Schuppan D, Becker J, Reichart P, Gelderblom
HR. Distribution of undulin, tenascin, and fibronectin in
the human periodontal ligament and cementum: com-
parative immunoelectron microscopy with ultra-thin cry-
osections. J Histochem Cytochem 1993: 41: 245251.
Nanci y Bosshardt
28
Potrebbero piacerti anche
- Anatomia Del PeriodontoDocumento33 pagineAnatomia Del PeriodontoMiSael PuiconNessuna valutazione finora
- Gingivitis CronicaDocumento47 pagineGingivitis CronicaAdriana Vargas100% (2)
- El Examen Periodontal Completo G C Armitage PDFDocumento12 pagineEl Examen Periodontal Completo G C Armitage PDFmarielNessuna valutazione finora
- Histopatologia PeriodonciaDocumento4 pagineHistopatologia PeriodonciaDelma Susana Trejo GurriaNessuna valutazione finora
- Marco Teorico Especifico y TratamientoDocumento11 pagineMarco Teorico Especifico y Tratamientoisrael tulcanazaNessuna valutazione finora
- Aseveraciones o ConstantesDocumento25 pagineAseveraciones o ConstantesFrancisco Javier Mata PerezNessuna valutazione finora
- Periodontología 2000Documento104 paginePeriodontología 2000Porras Zamudio LeninNessuna valutazione finora
- Fija I ResumenDocumento90 pagineFija I ResumenDAYSI ALEJANDRA GARCIA CALLESNessuna valutazione finora
- Cartilla MotivacionalDocumento10 pagineCartilla MotivacionalMaher Pajaro100% (1)
- Ancho o Espacio PeriodontalDocumento8 pagineAncho o Espacio PeriodontalS Andrea Hincapie PlataNessuna valutazione finora
- Enfoque Ortodoncico en El TX Multidisciplinario de PX Adultos. Relacion Con Periodoncia.Documento12 pagineEnfoque Ortodoncico en El TX Multidisciplinario de PX Adultos. Relacion Con Periodoncia.Leslie CMNessuna valutazione finora
- 7eecaf2d-2565-412f-8226-1b0d2f6796ca.docxDocumento6 pagine7eecaf2d-2565-412f-8226-1b0d2f6796ca.docxCarlosNessuna valutazione finora
- Unidad Dentogingival - BiologíaDocumento48 pagineUnidad Dentogingival - BiologíaMaria José HerreraNessuna valutazione finora
- Preparaciòn Dentaria Coronas CompletasDocumento5 paginePreparaciòn Dentaria Coronas CompletasMaria SanchezNessuna valutazione finora
- Tesis - Jonathan Loor FigueroaDocumento57 pagineTesis - Jonathan Loor FigueroaAlbertNessuna valutazione finora
- El Entorno OralDocumento2 pagineEl Entorno OralCeci Martinez50% (2)
- Bolsa PeriodontalDocumento23 pagineBolsa PeriodontalJeremy MontañezNessuna valutazione finora
- Métodos Control Tisular en La Toma de Impresión DefinitivaDocumento42 pagineMétodos Control Tisular en La Toma de Impresión DefinitivaKarla CruzNessuna valutazione finora
- PeriodontogramaDocumento3 paginePeriodontogramaYenifer Ccalli AlmonteNessuna valutazione finora
- Características Clinicas Del PeriodontoDocumento2 pagineCaracterísticas Clinicas Del PeriodontoMaria J Ordonez PerezNessuna valutazione finora
- Cirugia PeriodontalDocumento7 pagineCirugia PeriodontalaereenNessuna valutazione finora
- Bassetti 2016.en - EsDocumento19 pagineBassetti 2016.en - EsCarolina Dávila RamírezNessuna valutazione finora
- Mucogingival Conditions in The Natural Dentition - En.esDocumento10 pagineMucogingival Conditions in The Natural Dentition - En.eselizabeth grengerNessuna valutazione finora
- Concepto de PeriodonciaDocumento3 pagineConcepto de PeriodonciaJemelys Tejeda PlataNessuna valutazione finora
- UAP - Historia Clínica Integral Del Adulto 2010-2Documento14 pagineUAP - Historia Clínica Integral Del Adulto 2010-2Jhonatan Rodriguez AnguloNessuna valutazione finora
- Prueba de La Estructura MetalicaDocumento4 paginePrueba de La Estructura MetalicaStefany Bernal100% (1)
- Fenotipo GingivalDocumento12 pagineFenotipo GingivalAle Ardila OcampoNessuna valutazione finora
- Índices Epidemiológicos en OdontologíaDocumento16 pagineÍndices Epidemiológicos en OdontologíaAlejandraNessuna valutazione finora
- Cuadros 3Documento4 pagineCuadros 3Martha Piero Barone OgnibeneNessuna valutazione finora
- Resumenes Estomatologia B - 2014Documento103 pagineResumenes Estomatologia B - 2014Franco Botta100% (1)