Potrebbero piacerti anche
- Claudia Jones Nuclear TestingDocumento25 pagineClaudia Jones Nuclear TestingDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Continuity Change State of Process of Task ofDocumento1 paginaContinuity Change State of Process of Task ofDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Beta Anomaly An Ex-Ante Tail RiskDocumento104 pagineBeta Anomaly An Ex-Ante Tail RiskDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Theophilus Capital Against: Fisk Labor1Documento9 pagineTheophilus Capital Against: Fisk Labor1Daniel Lee Eisenberg Jacobs100% (1)
- Why The Euro Will Rival The Dollar PDFDocumento25 pagineWhy The Euro Will Rival The Dollar PDFDaniel Lee Eisenberg JacobsNessuna valutazione finora
- 1st International: Djacobs November 2020Documento5 pagine1st International: Djacobs November 2020Daniel Lee Eisenberg JacobsNessuna valutazione finora
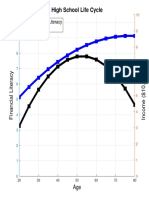
- High School Life Cycle: Financial Literacy IncomeDocumento1 paginaHigh School Life Cycle: Financial Literacy IncomeDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Conference Group For Central European History of The American Historical AssociationDocumento9 pagineConference Group For Central European History of The American Historical AssociationDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Karl Kautsky Republic and Social Democra PDFDocumento4 pagineKarl Kautsky Republic and Social Democra PDFDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Borel Sets PDFDocumento181 pagineBorel Sets PDFDaniel Lee Eisenberg Jacobs100% (1)
- Necessary and Sufficient Conditions For Dynamic OptimizationDocumento18 pagineNecessary and Sufficient Conditions For Dynamic OptimizationDaniel Lee Eisenberg JacobsNessuna valutazione finora
- The Saving Behavior of Public Vocational High School Students of Business and Management Program in Semarang SitiDocumento8 pagineThe Saving Behavior of Public Vocational High School Students of Business and Management Program in Semarang SitiDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Individualization of Robo-AdviceDocumento8 pagineIndividualization of Robo-AdviceDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Cover FERCDocumento1 paginaCover FERCDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Teach-In: Government of The People, by The People, For The PeopleDocumento24 pagineTeach-In: Government of The People, by The People, For The PeopleDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Gpebook PDFDocumento332 pagineGpebook PDFDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Bank Loan Loss ProvisioningDocumento17 pagineBank Loan Loss ProvisioningDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Simple BeamerDocumento25 pagineSimple BeamerDaniel Lee Eisenberg JacobsNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1091)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Biokimia I Carbohydrate: Dian Safitri F1C115017 Faculty of Math and Science Haluoleo UniversityDocumento14 pagineBiokimia I Carbohydrate: Dian Safitri F1C115017 Faculty of Math and Science Haluoleo UniversitytesyaNessuna valutazione finora
- Biochemistry - CarbohydratesDocumento40 pagineBiochemistry - CarbohydratesBalakrishnan RengeshNessuna valutazione finora
- BCH 201 - Ibbul - 2017Documento65 pagineBCH 201 - Ibbul - 2017Bello Ajetayo100% (1)
- Pharmacognosy PDFDocumento206 paginePharmacognosy PDFJunelle CostesNessuna valutazione finora
- Carbohydrates Part 4Documento46 pagineCarbohydrates Part 4Frnckk MnstrNessuna valutazione finora
- Azucares ReductoresDocumento6 pagineAzucares ReductoresKarla Edith Rodriguez NavaNessuna valutazione finora
- BIO F4 Chapter 4 Obj QuesttionDocumento3 pagineBIO F4 Chapter 4 Obj QuesttionIda KamalNessuna valutazione finora
- Karbohidrat: Gula, Pati & SeratDocumento56 pagineKarbohidrat: Gula, Pati & SeratAbdipramugaNessuna valutazione finora
- Science 10 4th Quarter ExamDocumento3 pagineScience 10 4th Quarter ExamCatherine Nadila33% (3)
- Learning Module General Biology 1Documento18 pagineLearning Module General Biology 1John Paull CuaNessuna valutazione finora
- DSE Bio Notes CH 1-4, 6, 20-28Documento212 pagineDSE Bio Notes CH 1-4, 6, 20-28ching100% (3)
- MONOSACCHARIDEDocumento2 pagineMONOSACCHARIDEPRINTDESK by Dan100% (1)
- Differential Behaviour of The Dinitrosalicylic Acid (DNS) ReagentDocumento3 pagineDifferential Behaviour of The Dinitrosalicylic Acid (DNS) Reagentmogn85Nessuna valutazione finora
- Defects of Carbohydrates Digestion & AbsorptionDocumento17 pagineDefects of Carbohydrates Digestion & AbsorptionMustafa KhandgawiNessuna valutazione finora
- Chemical Biology 03 Blood: Biomolecular Structure Lecture 3: 9/13/10Documento23 pagineChemical Biology 03 Blood: Biomolecular Structure Lecture 3: 9/13/10JuvvilyynGraceMahomatNessuna valutazione finora
- Nutri CarbsDocumento4 pagineNutri CarbsStacey GwenNessuna valutazione finora
- Handbook of General Animal NutritionDocumento303 pagineHandbook of General Animal Nutritionnaturalamir80% (5)
- UNIT4 Expt1Documento6 pagineUNIT4 Expt1Christian Franco RuizNessuna valutazione finora
- Alia Karbo PantulannDocumento23 pagineAlia Karbo PantulannRafidah AmaliaNessuna valutazione finora
- CarbohydrateDocumento116 pagineCarbohydrateManoj SigdelNessuna valutazione finora
- Microbio1 18Documento73 pagineMicrobio1 18Charles EspiloyNessuna valutazione finora
- Post Lab Experiment 3 Carbohydrates Group Work PDFDocumento8 paginePost Lab Experiment 3 Carbohydrates Group Work PDFJeff MarianoNessuna valutazione finora
- BIO 1401 Module Prof. C. Katongo-1Documento79 pagineBIO 1401 Module Prof. C. Katongo-1OliverNessuna valutazione finora
- Gen Bio Module 1Documento178 pagineGen Bio Module 1Ritney Jean AgpalasinNessuna valutazione finora
- Energy and CaloriesDocumento59 pagineEnergy and CaloriesKena SamuelNessuna valutazione finora
- Chapter 4 Bio NutrientsDocumento58 pagineChapter 4 Bio NutrientsNurain AiNessuna valutazione finora
- SWARNADocumento33 pagineSWARNASwetha KarunakaranNessuna valutazione finora
- G. Biology PPT Chapter 1-6new1Documento238 pagineG. Biology PPT Chapter 1-6new1Alemayehu MelkamuuNessuna valutazione finora
- Structure Function CarbohydratesDocumento28 pagineStructure Function Carbohydratespriya19866Nessuna valutazione finora
- CarbsDocumento34 pagineCarbsManendra PatwaNessuna valutazione finora