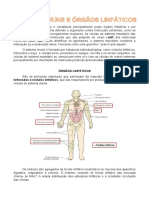
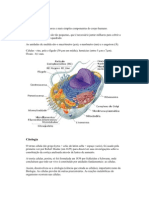
Potrebbero piacerti anche
- Bioquímica Clínica (Biomedicina) 1Documento27 pagineBioquímica Clínica (Biomedicina) 1Cinthya Carlene Terapias NaturaisNessuna valutazione finora
- Resumo & Guia De Estudo - Uma Elegante Defesa: A Extraordinária Nova Ciência Do Sistema ImunológicoDa EverandResumo & Guia De Estudo - Uma Elegante Defesa: A Extraordinária Nova Ciência Do Sistema ImunológicoNessuna valutazione finora
- Mecanismos de Agressão e Defesa PDFDocumento122 pagineMecanismos de Agressão e Defesa PDFSabrina FerreiraNessuna valutazione finora
- 9 Ef 13 DDocumento74 pagine9 Ef 13 DLaura ScottNessuna valutazione finora
- ImunologiaDocumento15 pagineImunologiaMarina Garcia100% (1)
- Resumo Do Livro Abbas de ImunologiaDocumento43 pagineResumo Do Livro Abbas de ImunologiaGustavo Tagliavini100% (4)
- Medresumo 2016 - Módulo 04 - Mecanismos de Agressao e Defesa (Mad) IIDocumento197 pagineMedresumo 2016 - Módulo 04 - Mecanismos de Agressao e Defesa (Mad) IIOsmair AlvesNessuna valutazione finora
- Digestao e Absorcao de LipidiosDocumento42 pagineDigestao e Absorcao de LipidiosLívia Born100% (1)
- Aula Introdução A ImunologiaDocumento53 pagineAula Introdução A ImunologiaFabrícia Lima FontesNessuna valutazione finora
- Estudo Dirigido BioquimicaDocumento5 pagineEstudo Dirigido Bioquimicaadelsonribeiro100% (4)
- Resumo Geral - AbbasDocumento24 pagineResumo Geral - AbbasVitor Hugo FerreiraNessuna valutazione finora
- Ficha de Trabalho Sobre BiomoléculasDocumento3 pagineFicha de Trabalho Sobre Biomoléculasspam mail100% (1)
- Ficha de Exercícios A Célula (VERSÃO CORRIGIDA) Ano Lectivo - 10º Ano Turma - DataDocumento10 pagineFicha de Exercícios A Célula (VERSÃO CORRIGIDA) Ano Lectivo - 10º Ano Turma - Datagelu1999Nessuna valutazione finora
- ImunologiaDocumento10 pagineImunologiaJanine Rafael100% (1)
- Aula MutaçõesDocumento24 pagineAula MutaçõesEloise DantasNessuna valutazione finora
- Propriedade Das Respostas ImunesDocumento5 paginePropriedade Das Respostas ImunesLara BorgesNessuna valutazione finora
- Resumo de Imunologia Completo-1Documento7 pagineResumo de Imunologia Completo-1Emanuelly Maria Da SilvaNessuna valutazione finora
- Resumo ImunologiaDocumento16 pagineResumo ImunologiaClaudiasenatore SenatoreNessuna valutazione finora
- Aula 1 - IntroduAAo A ImunologiaDocumento51 pagineAula 1 - IntroduAAo A ImunologiaLuizAlbuquerqueJr.Nessuna valutazione finora
- 6677 - Tas10 - 11Documento99 pagine6677 - Tas10 - 11leonor machadoNessuna valutazione finora
- 01 - Introdução Ao Sistema ImunológicoDocumento6 pagine01 - Introdução Ao Sistema ImunológicoDiego FonsecaNessuna valutazione finora
- Resumos de ImunologiaDocumento8 pagineResumos de ImunologiaJuliana LimaNessuna valutazione finora
- Resumo Histologia Do Sistema Imune e Órgãos LinfáticosDocumento13 pagineResumo Histologia Do Sistema Imune e Órgãos LinfáticosAlexandre AugustusNessuna valutazione finora
- Cap 1 - Propriedades Gerais Das Respostas ImunesDocumento5 pagineCap 1 - Propriedades Gerais Das Respostas ImunesMatheusNessuna valutazione finora
- TrabimunoDocumento10 pagineTrabimunoMICHELINENessuna valutazione finora
- ESTUDO DIRIGIDO AV2 MICRO E IMUNO 2021 2s-1 ComrespostaDocumento4 pagineESTUDO DIRIGIDO AV2 MICRO E IMUNO 2021 2s-1 ComrespostaDaniel FerreiraNessuna valutazione finora
- Imunologia Aula 1Documento5 pagineImunologia Aula 1RhuanLopesNessuna valutazione finora
- ImunologiaDocumento26 pagineImunologiaThayane OliveiraNessuna valutazione finora
- Cárie Dentária Introdução IDocumento207 pagineCárie Dentária Introdução ICibele Correia100% (1)
- Questões para Estudo de ImunologiaDocumento9 pagineQuestões para Estudo de ImunologiaauroeagNessuna valutazione finora
- Resumo Imunologia - Principais Conceitos Do Capítulos 01 e 05 ABBASDocumento9 pagineResumo Imunologia - Principais Conceitos Do Capítulos 01 e 05 ABBASMatheus RodriguesNessuna valutazione finora
- Imunologia IDocumento3 pagineImunologia Ifeliaraujo16Nessuna valutazione finora
- Introdução A Imunologia - Aula 1Documento7 pagineIntrodução A Imunologia - Aula 1manpa.ouroazulNessuna valutazione finora
- Pós Aulas Encontro 9 Ao Encontro 15 - MicrobiologiaDocumento11 paginePós Aulas Encontro 9 Ao Encontro 15 - MicrobiologiaAssociação Jardins Roma 34 -3214-2314Nessuna valutazione finora
- Imunidade Adaptativa - 2023Documento33 pagineImunidade Adaptativa - 2023Carolayne Souza PintoNessuna valutazione finora
- Avaliacao Imunologia FernandaDocumento6 pagineAvaliacao Imunologia FernandaFernanda SampaioNessuna valutazione finora
- Estudo Dirigido ImunologiaDocumento7 pagineEstudo Dirigido ImunologiaLeticiaNessuna valutazione finora
- Cap 1, ABBASDocumento5 pagineCap 1, ABBASMedicina.aNessuna valutazione finora
- Resumo de Imunologia - Barbara Ribeiro XLVIIIDocumento16 pagineResumo de Imunologia - Barbara Ribeiro XLVIIINonato Oliveira Carvalho Junior100% (1)
- Apostila Básica de IMUNOLOGIA VETERINÁRIADocumento16 pagineApostila Básica de IMUNOLOGIA VETERINÁRIALorena BandeiraNessuna valutazione finora
- Imunidade e Controlo de DoençasDocumento49 pagineImunidade e Controlo de DoençasLourenço_290% (10)
- ImunoguiaDocumento101 pagineImunoguiaMateus Barbosa RodriguesNessuna valutazione finora
- RPH ImunologiaDocumento24 pagineRPH ImunologiarotacomandoNessuna valutazione finora
- Tutorial 1 - ProliferaçãoDocumento16 pagineTutorial 1 - ProliferaçãoStephany VitóriaNessuna valutazione finora
- Resumo Imunologia VETERINÁRIADocumento8 pagineResumo Imunologia VETERINÁRIAGiovanna CherubiniNessuna valutazione finora
- Sistema ImunológicoDocumento17 pagineSistema Imunológicojoyce santosNessuna valutazione finora
- 2º Aula - Imunidade Adaptativa PDFDocumento4 pagine2º Aula - Imunidade Adaptativa PDFLeonardo Siqueira RamosNessuna valutazione finora
- Final 7906Documento8 pagineFinal 7906AnaRafaelaSilvaNessuna valutazione finora
- Prints e Resumos ImunologiaDocumento5 paginePrints e Resumos ImunologiaAna LídiaNessuna valutazione finora
- Fichamento Dinâmica Da Resposta Imune ImunologiaDocumento1 paginaFichamento Dinâmica Da Resposta Imune ImunologiaJulia Lopes MouraNessuna valutazione finora
- Imunopatologia Aula 2Documento36 pagineImunopatologia Aula 2adilsonjacinto19945Nessuna valutazione finora
- Sistem AimuneDocumento57 pagineSistem AimunepriscilaqueengomesfNessuna valutazione finora
- ImunologiaDocumento9 pagineImunologiaAlexia LimaNessuna valutazione finora
- ImunologiaDocumento284 pagineImunologiaAntonio LopezNessuna valutazione finora
- Resumo Imunologia P1Documento18 pagineResumo Imunologia P1exameslaboratoriais2013Nessuna valutazione finora
- Estudo Dirigido de Imunologia para P1Documento2 pagineEstudo Dirigido de Imunologia para P1Vanessa Suely Campos100% (2)
- Células e Órgãos Do Sistema ImuneDocumento10 pagineCélulas e Órgãos Do Sistema ImuneRicardo RamsesNessuna valutazione finora
- Imuno - Células e OrgãosDocumento5 pagineImuno - Células e OrgãosKarin TreichelNessuna valutazione finora
- Ficha Resição Biologia Funcional - Sistema ImuneDocumento18 pagineFicha Resição Biologia Funcional - Sistema ImuneDiogo Botelho Medeiros BrilhanteNessuna valutazione finora
- Atividade 500 Pontos 29 10 2020Documento9 pagineAtividade 500 Pontos 29 10 2020Luuh MacieiraNessuna valutazione finora
- Livro-Texto - Unidade IIDocumento34 pagineLivro-Texto - Unidade IIRafael CedroNessuna valutazione finora
- Aula 5 - Fisiologia Do Sistema Imune (Atualização)Documento12 pagineAula 5 - Fisiologia Do Sistema Imune (Atualização)Lázaro Victor Santos Mendonça100% (1)
- Imunologia - Estudo Dirigido 02Documento3 pagineImunologia - Estudo Dirigido 02Dan LionNessuna valutazione finora
- Imunologia ResumoDocumento10 pagineImunologia ResumocdromcreativeNessuna valutazione finora
- 2 - Revisão Da ImunologiaDocumento7 pagine2 - Revisão Da ImunologiaPaulo Fernando Cardoso0% (1)
- Estudo Dirigido BioquimicaDocumento2 pagineEstudo Dirigido BioquimicaCiroNessuna valutazione finora
- Resumo Respiração Aeróbica e AnaeróbicaDocumento11 pagineResumo Respiração Aeróbica e AnaeróbicaDanielNessuna valutazione finora
- Apostila VitaminasDocumento7 pagineApostila VitaminasMaria Emilia CoimbraNessuna valutazione finora
- AULA 18 Sebenta de BactereologiaDocumento22 pagineAULA 18 Sebenta de Bactereologiaapi-3826744100% (2)
- CEDERJ Biologia Celular I - Gabarito 21 A 30Documento9 pagineCEDERJ Biologia Celular I - Gabarito 21 A 30api-3800070Nessuna valutazione finora
- Ácidos NucleicosDocumento14 pagineÁcidos NucleicosMilene Borges MendesNessuna valutazione finora
- Natureza e Estrutura Do Material Genético - BM 2007Documento62 pagineNatureza e Estrutura Do Material Genético - BM 2007Alexandra SalvadoNessuna valutazione finora
- Nivel Celular 10 ClasseDocumento10 pagineNivel Celular 10 ClasseLunguequi Júlio De Jesus MuzolaNessuna valutazione finora
- NUTRICÃO DE NÃO RUMINATES - Material de EstudoDocumento53 pagineNUTRICÃO DE NÃO RUMINATES - Material de EstudoSilva, M.S.J.Nessuna valutazione finora
- Escola Secundária Pedro Nune2Documento10 pagineEscola Secundária Pedro Nune2António LucasNessuna valutazione finora
- Acidos NucleicosDocumento18 pagineAcidos NucleicoslarissaNessuna valutazione finora
- ANTINEOPLÁSICOSDocumento18 pagineANTINEOPLÁSICOSMichelly Gonçalves KrapienisNessuna valutazione finora
- Teste2 Nov2015 V1Documento9 pagineTeste2 Nov2015 V1José BaptistaNessuna valutazione finora
- LipídiosDocumento2 pagineLipídiosLais MarquesNessuna valutazione finora
- Introducao Ao Estudo Da GastronomiaDocumento17 pagineIntroducao Ao Estudo Da GastronomiaAngel Tuga100% (1)
- Aula On-Line 6 EnzimasDocumento47 pagineAula On-Line 6 EnzimasPaula Polvora VictoriaNessuna valutazione finora
- Célula AnimalDocumento6 pagineCélula AnimalFelipe MeloNessuna valutazione finora
- Proteínas Na Nutrição de Cães e GatosDocumento14 pagineProteínas Na Nutrição de Cães e GatosDébora SakiyamaNessuna valutazione finora
- 2CES FE CFB Biologia 10Documento26 pagine2CES FE CFB Biologia 10Manuel António Jamba JambaNessuna valutazione finora
- Morosil 1Documento2 pagineMorosil 1AdrianaNessuna valutazione finora
- Seminario de Bioquimica OfcDocumento57 pagineSeminario de Bioquimica OfcDébora CarvalhoNessuna valutazione finora
- Teste 1 - 11 AnoDocumento8 pagineTeste 1 - 11 Anomjoao193Nessuna valutazione finora
- 5 mch9 DigestivoDocumento39 pagine5 mch9 DigestivoevanunesNessuna valutazione finora