Potrebbero piacerti anche
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Da EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Nessuna valutazione finora
- Biologia Apunte DefinitivoDocumento108 pagineBiologia Apunte DefinitivoOlivia CanelaNessuna valutazione finora
- Biologia Celular Banco de Preguntas FinalDocumento14 pagineBiologia Celular Banco de Preguntas Finalcharit martinez ñomaNessuna valutazione finora
- Transporte Por PermeabilidadDocumento10 pagineTransporte Por PermeabilidadaaliyahNessuna valutazione finora
- MACHETE2CAMPUS2020. Biomembranas... Docx PDFDocumento22 pagineMACHETE2CAMPUS2020. Biomembranas... Docx PDFMagentaIdeasImpresasNessuna valutazione finora
- Sesión 04 - Membrana Celular - Estructura y ComposiciónDocumento24 pagineSesión 04 - Membrana Celular - Estructura y ComposiciónKaaterin Valeria Mestanza MacahuachiNessuna valutazione finora
- Tema 7. Las Envolturas CelularesDocumento12 pagineTema 7. Las Envolturas CelularesMarco MatellánNessuna valutazione finora
- CélulaDocumento12 pagineCélulabalumbishNessuna valutazione finora
- Cuestionario 1Documento8 pagineCuestionario 1Berenice S. OleaNessuna valutazione finora
- Apuntes Biologia 2Documento23 pagineApuntes Biologia 2mbelenruhlNessuna valutazione finora
- En Volturas Celulares 2º BachDocumento5 pagineEn Volturas Celulares 2º BachMónica Fernández SantosNessuna valutazione finora
- Seminario 1 (Práctica) - Informe PDFDocumento17 pagineSeminario 1 (Práctica) - Informe PDFFernanda Pita ReyesNessuna valutazione finora
- Fisiopatología y Membrana CelularDocumento49 pagineFisiopatología y Membrana CelularJazmin Sanchez Sanchez100% (2)
- Tema 2Documento22 pagineTema 2Endika ValladolidNessuna valutazione finora
- Membrana Plasmatica - BioquimicaDocumento7 pagineMembrana Plasmatica - BioquimicaNaomi Nathaly Castañeda AnampaNessuna valutazione finora
- EndomembranasDocumento5 pagineEndomembranasn59vjk68wpNessuna valutazione finora
- RESUMENDocumento6 pagineRESUMENKimMartinTNessuna valutazione finora
- Fisiología 1Documento19 pagineFisiología 1Rocío TempranoNessuna valutazione finora
- Resumen Cap 13 de CooperDocumento5 pagineResumen Cap 13 de CooperLeao Cornejo VeraNessuna valutazione finora
- Transporte de SustanciasDocumento77 pagineTransporte de SustanciasClaudia Vargas AndinoNessuna valutazione finora
- Transporte A Traves de La MembranaDocumento34 pagineTransporte A Traves de La MembranaMaria Jose RO100% (1)
- Membranas BiologicasDocumento15 pagineMembranas BiologicasCelia Lizbeth Vallejos VelezNessuna valutazione finora
- Membrana y Transporte PDFDocumento48 pagineMembrana y Transporte PDFMarielos VelásquezNessuna valutazione finora
- Guia de BioDocumento9 pagineGuia de BioMelina HernandezNessuna valutazione finora
- Membrana CelularDocumento4 pagineMembrana CelularMichelle Estefanía Arredondo ArmasNessuna valutazione finora
- Guia ToraxDocumento17 pagineGuia Toraxignacia arteagaNessuna valutazione finora
- Retículo EndoplasmáticoDocumento5 pagineRetículo EndoplasmáticoKarla Peláez GalánNessuna valutazione finora
- Guia PDF Transporte de MembranaDocumento5 pagineGuia PDF Transporte de Membrananicole_sierraangel100% (1)
- Osmosis y Transporte CelularDocumento6 pagineOsmosis y Transporte Celularkyomich100% (1)
- Transporte CelularDocumento8 pagineTransporte CelularkathericNessuna valutazione finora
- Membrana Plasmática. Modelo Mosaico FluidoDocumento3 pagineMembrana Plasmática. Modelo Mosaico FluidoJavier Rama GonzálezNessuna valutazione finora
- T7 La Membrana CelularDocumento7 pagineT7 La Membrana Celularaitana.roso06Nessuna valutazione finora
- Citologia BasicaDocumento33 pagineCitologia BasicaJhon Salas Mallqui0% (1)
- Funciones de La Membrana CelularDocumento12 pagineFunciones de La Membrana CelularGiovannaNessuna valutazione finora
- Vesículas de Transporte o de TransiciónDocumento4 pagineVesículas de Transporte o de TransiciónÁlex Willan Acuña SilvaNessuna valutazione finora
- El OvuloDocumento8 pagineEl OvuloMeimis TorregrozaNessuna valutazione finora
- RESÚMEN Temas de Tercero Biología SecundariaDocumento21 pagineRESÚMEN Temas de Tercero Biología SecundariaFlorencia UrristiNessuna valutazione finora
- Repaso 2Documento7 pagineRepaso 2María Isabel OrdenesNessuna valutazione finora
- Tema 4Documento6 pagineTema 4Erika Luna JiménezNessuna valutazione finora
- Transporte CelularDocumento8 pagineTransporte CelularCesar Manuel Morales AbreuNessuna valutazione finora
- Test de BiologiaDocumento11 pagineTest de BiologiaCamila G. GutiérrezNessuna valutazione finora
- Biología Molecular de Los Transportadores de GlucosaDocumento38 pagineBiología Molecular de Los Transportadores de GlucosaCecilia Marín ZambranoNessuna valutazione finora
- Transporte de MembranaDocumento13 pagineTransporte de MembranaLalaNessuna valutazione finora
- Guía Transporte Pasivo y ActivoDocumento7 pagineGuía Transporte Pasivo y ActivoMARIA PIA VALENZUELA JARANessuna valutazione finora
- La Membrana CelularDocumento68 pagineLa Membrana CelularMaría Ángel DoriaNessuna valutazione finora
- Capitulo 7 Estructura y Funcion de La MembranaDocumento4 pagineCapitulo 7 Estructura y Funcion de La MembranaRicardo PosadasNessuna valutazione finora
- Fisiología de La Membrana MGT FLUKERDocumento37 pagineFisiología de La Membrana MGT FLUKER023100894hNessuna valutazione finora
- BIOLOGIADocumento7 pagineBIOLOGIAPatty RamirezNessuna valutazione finora
- Tema 2 - Membrana Celular y CitoplasmaDocumento11 pagineTema 2 - Membrana Celular y CitoplasmaALETEPNessuna valutazione finora
- Biología Re y GolgiDocumento6 pagineBiología Re y GolgiNabila InsfranNessuna valutazione finora
- Membranas y TransporteDocumento5 pagineMembranas y TransporteRojo RubiNessuna valutazione finora
- Membranas y CélulaDocumento27 pagineMembranas y CélulaodjibwaNessuna valutazione finora
- Fisiología de La Membrana CelularDocumento6 pagineFisiología de La Membrana CelularMax ChafloqueNessuna valutazione finora
- Transporte CelularDocumento49 pagineTransporte CelularOrlando Arian SánchezNessuna valutazione finora
- Membrana CelularDocumento10 pagineMembrana Celularapi-3762846100% (6)
- Histologia Guía de EstudioDocumento37 pagineHistologia Guía de EstudioIdelberto BenitezNessuna valutazione finora
- Biologia Molecular de Los Transport Adores de GlucosaDocumento11 pagineBiologia Molecular de Los Transport Adores de GlucosaFabiola MastromatteoNessuna valutazione finora
- Capitulo VI. Transporte Vesicular de Las Rutas SecretorasDocumento12 pagineCapitulo VI. Transporte Vesicular de Las Rutas SecretorasCamila Paulina Aravena SotoNessuna valutazione finora
- La Membrana Celular.Documento76 pagineLa Membrana Celular.F1101Nessuna valutazione finora
- Membrana PlasmáticaDocumento12 pagineMembrana PlasmáticavinicioNessuna valutazione finora
- AcvDocumento113 pagineAcvLaura García HenaoNessuna valutazione finora
- Surviving Sepsis Campaign 2016 Guidelines Presentation Final EspañolDocumento54 pagineSurviving Sepsis Campaign 2016 Guidelines Presentation Final EspañolLaura García Henao100% (1)
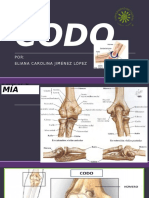
- CODODocumento48 pagineCODOLaura García HenaoNessuna valutazione finora
- Patología Laríngea en PediatríaDocumento76 paginePatología Laríngea en PediatríaLaura García HenaoNessuna valutazione finora
- Caso Clínico Geriatría 1Documento62 pagineCaso Clínico Geriatría 1Laura García HenaoNessuna valutazione finora
- Tamizaje CA de Cuello Uterino FinalDocumento17 pagineTamizaje CA de Cuello Uterino FinalLaura García HenaoNessuna valutazione finora
- Manual Merk de Signos y SintomasDocumento642 pagineManual Merk de Signos y SintomasJahirAleaxanderJibajaCedilloNessuna valutazione finora
- Medicamentos Biotecnológicos ListoDocumento19 pagineMedicamentos Biotecnológicos ListoLaura García HenaoNessuna valutazione finora
- Exposición DiarreaDocumento23 pagineExposición DiarreaLaura García HenaoNessuna valutazione finora
- Dermatología Práctica PDFDocumento102 pagineDermatología Práctica PDFLaura García Henao100% (5)
- Aewebpoldiae34761124 PDFDocumento11 pagineAewebpoldiae34761124 PDFsergioNessuna valutazione finora
- Val 2005 CatalogoDocumento50 pagineVal 2005 CatalogoAlvaro Sebastián CamachoNessuna valutazione finora
- Izaje de CargasDocumento91 pagineIzaje de Cargasmary paredesNessuna valutazione finora
- Transporte MultimodalDocumento23 pagineTransporte MultimodalSamantha BaezNessuna valutazione finora
- Infografia Seguridad Vial y Manejo Defensivo CHDocumento6 pagineInfografia Seguridad Vial y Manejo Defensivo CHFreddy George Gonzales MendietaNessuna valutazione finora
- Ficha DHP458Documento1 paginaFicha DHP458Vanesa SanzNessuna valutazione finora
- Linea de Tiempo - Ferrocarril en ColombiaDocumento2 pagineLinea de Tiempo - Ferrocarril en ColombiaNicolás Ticora67% (3)
- Estabilidad de TaludesDocumento19 pagineEstabilidad de Taludesrodolfo201267% (6)
- Tarea Academica 1Documento11 pagineTarea Academica 1Kataleya PerezNessuna valutazione finora
- A3 - Taller3Documento3 pagineA3 - Taller3giselaNessuna valutazione finora
- Calculo de Estabilidad BarcazaDocumento14 pagineCalculo de Estabilidad Barcazarene_poma4426Nessuna valutazione finora
- K-Cc-147c-Hse-Jsa-051 - RH 20.04.18Documento56 pagineK-Cc-147c-Hse-Jsa-051 - RH 20.04.18diego recabarrenNessuna valutazione finora
- BateriaDocumento62 pagineBateriaFeanco mohroNessuna valutazione finora
- Costos Unitarios EstructurasDocumento42 pagineCostos Unitarios EstructurasAldo Suárez ContrerasNessuna valutazione finora
- Principales Ríos de La Región ArequipaDocumento5 paginePrincipales Ríos de La Región ArequipaFernando Barra0% (1)
- Plan de Seguridad Vial 2018Documento33 paginePlan de Seguridad Vial 2018consorcio de transportistas san rafaelNessuna valutazione finora
- 58d421f7a046d PDFDocumento266 pagine58d421f7a046d PDFCerasani RoccoNessuna valutazione finora
- BarbanegraDocumento41 pagineBarbanegraDaniel Sierra MellaNessuna valutazione finora
- Especificaciones TecnicasDocumento22 pagineEspecificaciones TecnicasBrayan Antony Zamalloa ParoNessuna valutazione finora
- Definición de Mezcla AsfálticaDocumento14 pagineDefinición de Mezcla AsfálticaDaniel PerezNessuna valutazione finora
- Cronograma IcbfDocumento9 pagineCronograma IcbfEfrain CampoNessuna valutazione finora
- Primera Vez en El AeropuertoDocumento8 paginePrimera Vez en El AeropuertoHector GerardoNessuna valutazione finora
- Manual Mantenimiento (Jefe de Operaciones)Documento50 pagineManual Mantenimiento (Jefe de Operaciones)dionelcito67% (3)
- Leyenda Instalacion de DesagüeDocumento1 paginaLeyenda Instalacion de DesagüeLUIS GUTIERREZNessuna valutazione finora
- Actividad - Lección 1Documento4 pagineActividad - Lección 1Francis Paul Correa Giron100% (1)
- 61 HRC 1000 VDocumento1 pagina61 HRC 1000 VMateo Cab PirataNessuna valutazione finora
- MM-F-10 Inspección Bulldozer V05Documento2 pagineMM-F-10 Inspección Bulldozer V05Alfonso PrietoNessuna valutazione finora
- Trabajos en Altura C.I.A.S.Documento98 pagineTrabajos en Altura C.I.A.S.edy100% (1)
- Control de Multitudes VII PNDocumento40 pagineControl de Multitudes VII PNAleex RG100% (2)
- Cuadernillo IPV2Documento15 pagineCuadernillo IPV2KEVIN JOS POTES POTESNessuna valutazione finora