Potrebbero piacerti anche
- Long Non-coding RNA: The Dark Side of the GenomeDa EverandLong Non-coding RNA: The Dark Side of the GenomeNessuna valutazione finora
- The Role of WNT Signaling in Axis Development and Stem Cell Regulation in CnidariansDocumento6 pagineThe Role of WNT Signaling in Axis Development and Stem Cell Regulation in CnidariansEliasNessuna valutazione finora
- Wnt Signaling in Development and Disease: Molecular Mechanisms and Biological FunctionsDa EverandWnt Signaling in Development and Disease: Molecular Mechanisms and Biological FunctionsNessuna valutazione finora
- Dissertation WNT SignalwegDocumento4 pagineDissertation WNT SignalwegBestPaperWritingServiceReviewsSingapore100% (1)
- WNT Pathway - Zhan 2017Documento13 pagineWNT Pathway - Zhan 2017ALEXANDRA ANTONIETA URRUTIA ZEGARRANessuna valutazione finora
- Dev 146589Documento8 pagineDev 146589Zeynep YazıcıoğluNessuna valutazione finora
- Genes Dev.-2014-De Lau-305-16Documento13 pagineGenes Dev.-2014-De Lau-305-16Karthik AripakaNessuna valutazione finora
- Ferrer Vaquer 2010 A Sensitive and Bright Single-Cell Resolution Live Imaging Reporter of WNT Beta Catenin Signalling in MouseDocumento18 pagineFerrer Vaquer 2010 A Sensitive and Bright Single-Cell Resolution Live Imaging Reporter of WNT Beta Catenin Signalling in MouseAlejandro Farrera HernándezNessuna valutazione finora
- Regulated Wnt/Beta-Catenin Signaling Sustains Adult Spermatogenesis in MiceDocumento12 pagineRegulated Wnt/Beta-Catenin Signaling Sustains Adult Spermatogenesis in MiceamarNessuna valutazione finora
- A Single-Cell Atlas of Plasmodium Falciparum Transmission Through The MosquitoDocumento13 pagineA Single-Cell Atlas of Plasmodium Falciparum Transmission Through The MosquitoPatricia GomesNessuna valutazione finora
- Research Progression PresentDocumento40 pagineResearch Progression Presentt13ersNessuna valutazione finora
- Lincoln Stein - Genome Annotation: From Sequence To BiologyDocumento13 pagineLincoln Stein - Genome Annotation: From Sequence To BiologyYopghm698Nessuna valutazione finora
- Insertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal TumorigenesisDocumento10 pagineInsertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal Tumorigenesistalha saleemNessuna valutazione finora
- Anita Hall - WntsDocumento6 pagineAnita Hall - WntsMiles NsgNessuna valutazione finora
- The Cell Biology of Tobacco Mosaic Virus Replication and MovementDocumento10 pagineThe Cell Biology of Tobacco Mosaic Virus Replication and MovementKarim_pvdNessuna valutazione finora
- Assembly of Protein Complexes Restricts DiffusionDocumento15 pagineAssembly of Protein Complexes Restricts DiffusionRifqi Fathul ArroisiNessuna valutazione finora
- NIH Public Access: Author ManuscriptDocumento33 pagineNIH Public Access: Author ManuscriptHari PriyaNessuna valutazione finora
- Y-Linker 2Documento5 pagineY-Linker 2hadeelNessuna valutazione finora
- Huntingtin PesquisaDocumento2 pagineHuntingtin Pesquisamikey12345452863879Nessuna valutazione finora
- tmp3831 TMPDocumento3 paginetmp3831 TMPFrontiersNessuna valutazione finora
- Special Review Series - Gene Manipulation and Integrative PhysiologyDocumento13 pagineSpecial Review Series - Gene Manipulation and Integrative PhysiologyAnunay JoshiNessuna valutazione finora
- Journal Developmental BioDocumento11 pagineJournal Developmental Biodod_nurNessuna valutazione finora
- BCH4125 2022 - Lecture4 5 Jan20 25Documento61 pagineBCH4125 2022 - Lecture4 5 Jan20 25Fatima AkbariNessuna valutazione finora
- Touch NeuronsDocumento7 pagineTouch NeuronscesarjbcNessuna valutazione finora
- Advances in Esophageal Cancer A New Perspective On Pathogenes - 2018 - Cancer LDocumento8 pagineAdvances in Esophageal Cancer A New Perspective On Pathogenes - 2018 - Cancer LFlorin AchimNessuna valutazione finora
- Nihms684028 PDFDocumento26 pagineNihms684028 PDFSeptebi Eko PutraNessuna valutazione finora
- My TopicDocumento9 pagineMy TopicShailendra YadavNessuna valutazione finora
- C Elegans DissertationDocumento7 pagineC Elegans DissertationBuySchoolPapersOnlineSingapore100% (1)
- Crosstalk Between WNT and Bone Morphogenic Protein Signaling - A Turbulent RelationshipDocumento18 pagineCrosstalk Between WNT and Bone Morphogenic Protein Signaling - A Turbulent Relationshipaaki2627Nessuna valutazione finora
- Liman PNAS99 VNOchannelDocumento6 pagineLiman PNAS99 VNOchannelJuan David Avila RinconNessuna valutazione finora
- Liman PNAS99 VNOchannelDocumento6 pagineLiman PNAS99 VNOchannelJuan David Avila RinconNessuna valutazione finora
- Liman PNAS99 VNOchannelDocumento6 pagineLiman PNAS99 VNOchannelJuan David Avila RinconNessuna valutazione finora
- Secretion and Extracellular Space Travel of WNT ProteinsDocumento6 pagineSecretion and Extracellular Space Travel of WNT ProteinsEduardo RodríguezNessuna valutazione finora
- Tenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionDocumento12 pagineTenascin-C Promotes Tumor Cell Migration and Metastasis Through Integrin A9b1-Mediated YAP InhibitionSayda DhaouadiNessuna valutazione finora
- Leptin 1Documento9 pagineLeptin 1vestibuleNessuna valutazione finora
- GAMETOGENESISDocumento44 pagineGAMETOGENESISMary Eunice Elizabeth YraNessuna valutazione finora
- Anastas 2015Documento46 pagineAnastas 2015Mehedi HossainNessuna valutazione finora
- Inbound 8785119957122048194Documento15 pagineInbound 8785119957122048194MUHAMMAD ISHAQNessuna valutazione finora
- The Use of Zebrafish To ReviewDocumento13 pagineThe Use of Zebrafish To ReviewKarthik ShettyNessuna valutazione finora
- BMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in MiceDocumento14 pagineBMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in Micefather45Nessuna valutazione finora
- Wnt/ β-catenin signalling: function, biological mechanisms, and therapeutic opportunitiesDocumento23 pagineWnt/ β-catenin signalling: function, biological mechanisms, and therapeutic opportunitiesOriginZeroNessuna valutazione finora
- Dev 201531Documento18 pagineDev 201531nzumana258Nessuna valutazione finora
- A Last WordDocumento5 pagineA Last WordBruna Klippel FerreiraNessuna valutazione finora
- ARTICULO 1cell Signallingcornell2005Documento4 pagineARTICULO 1cell Signallingcornell2005Carmen MaRiNessuna valutazione finora
- Ppat 1007985Documento23 paginePpat 1007985bilou hertNessuna valutazione finora
- WNT Signaling in TheDocumento29 pagineWNT Signaling in ThehappyanylandNessuna valutazione finora
- BCH4125 2022 - Lecture6 Jan27Documento27 pagineBCH4125 2022 - Lecture6 Jan27Fatima AkbariNessuna valutazione finora
- 2010 - The Zebrafish As A Model Organism For The Study of Apoptosis PDFDocumento19 pagine2010 - The Zebrafish As A Model Organism For The Study of Apoptosis PDFSebastián PiernagordaNessuna valutazione finora
- Neural Development in HumansDocumento14 pagineNeural Development in Humanssandeshi1Nessuna valutazione finora
- Shekinah1 PDFDocumento10 pagineShekinah1 PDFShekinah Andrea DumlaoNessuna valutazione finora
- Art:10.1007/s11738 010 0538 2 PDFDocumento12 pagineArt:10.1007/s11738 010 0538 2 PDFPedro ConfortNessuna valutazione finora
- Emb Oj 2012146 ADocumento15 pagineEmb Oj 2012146 Abiotech_vidhyaNessuna valutazione finora
- Transgenic AnimalsDocumento12 pagineTransgenic AnimalsAbhimanyu JainNessuna valutazione finora
- Transposon-Activated POU5F1B Promotes Colorectal Cancer Growth and MetastasisDocumento17 pagineTransposon-Activated POU5F1B Promotes Colorectal Cancer Growth and MetastasisJennifer WuNessuna valutazione finora
- Microorganisms v11 I03 20230622Documento15 pagineMicroorganisms v11 I03 20230622Editor IJDMNessuna valutazione finora
- Plant Virus InteractionDocumento7 paginePlant Virus InteractionShubham SutarNessuna valutazione finora
- Cores of Reproducibility in Physiology: CORP: Using Transgenic Mice To Study Skeletal Muscle Physiology - PMCDocumento26 pagineCores of Reproducibility in Physiology: CORP: Using Transgenic Mice To Study Skeletal Muscle Physiology - PMCd2cc8hbcjcNessuna valutazione finora
- Biochemical and Biophysical Research CommunicationsDocumento6 pagineBiochemical and Biophysical Research CommunicationsSergeat18BNessuna valutazione finora
- Transgenic Animal FarmDocumento20 pagineTransgenic Animal FarmRichard WooliteNessuna valutazione finora
- WNT Signaling Multiple PathwaysDocumento6 pagineWNT Signaling Multiple PathwaysMayerlis ArrietaNessuna valutazione finora
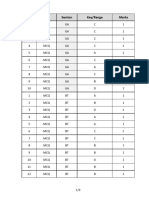
- Q.No. Type Section Key/Range MarksDocumento3 pagineQ.No. Type Section Key/Range Marksbiotech_vidhyaNessuna valutazione finora
- Buffer Preparation PDFDocumento6 pagineBuffer Preparation PDFbiotech_vidhyaNessuna valutazione finora
- BT 2019Documento13 pagineBT 2019biotech_vidhyaNessuna valutazione finora
- Troubleshooting SDS-PAGE 1Documento3 pagineTroubleshooting SDS-PAGE 1biotech_vidhyaNessuna valutazione finora
- Polymerase Chain Reaction (PCR)Documento3 paginePolymerase Chain Reaction (PCR)biotech_vidhyaNessuna valutazione finora
- TNPSC Group 1 Prelim Book List PDFDocumento2 pagineTNPSC Group 1 Prelim Book List PDFbiotech_vidhyaNessuna valutazione finora
- Whole Cell ExtractDocumento1 paginaWhole Cell Extractbiotech_vidhyaNessuna valutazione finora
- Nuclear ExtractsDocumento2 pagineNuclear Extractsbiotech_vidhyaNessuna valutazione finora
- Ies 17 Set A Me Q ADocumento67 pagineIes 17 Set A Me Q Abiotech_vidhyaNessuna valutazione finora
- Mechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010Documento20 pagineMechanical Engineering Code No. 14: Combined Competitive (Preliminary) Examination, 2010biotech_vidhyaNessuna valutazione finora
- Isa BusDocumento30 pagineIsa Busbiotech_vidhya100% (1)
- Teachers Recruitment Board: 1. Important DatesDocumento13 pagineTeachers Recruitment Board: 1. Important Datesbiotech_vidhyaNessuna valutazione finora
- Befcv List PDFDocumento22 pagineBefcv List PDFbiotech_vidhyaNessuna valutazione finora
- 1 TolerancesDocumento1 pagina1 Tolerancesbiotech_vidhyaNessuna valutazione finora
- Lec09 Three PhaseDocumento84 pagineLec09 Three Phasebiotech_vidhyaNessuna valutazione finora
- Converting Saltwater Into Drinkwater As A Solution For Global Crisis in A Fresh Water Needs Using Green TechnologyDocumento4 pagineConverting Saltwater Into Drinkwater As A Solution For Global Crisis in A Fresh Water Needs Using Green TechnologyNurrahmah AzizahNessuna valutazione finora
- Surface Treatments For MG Alloys in Aerospace and DefenceDocumento16 pagineSurface Treatments For MG Alloys in Aerospace and DefenceJinsoo KimNessuna valutazione finora
- In Uence of Pre-Gelatinised Maize Starch On The Rheology, Microstructure and Processing of Imitation CheeseDocumento8 pagineIn Uence of Pre-Gelatinised Maize Starch On The Rheology, Microstructure and Processing of Imitation CheeseLina1929Nessuna valutazione finora
- IgsDocumento6 pagineIgsmrjn1367Nessuna valutazione finora
- Example CoagulFloccu1Documento3 pagineExample CoagulFloccu1Adarsh Agrawal100% (3)
- CSIR Physics Sciences Solved December 2012Documento43 pagineCSIR Physics Sciences Solved December 2012kesavavamsikrishnaNessuna valutazione finora
- Ana Bruker Scion TQ Spec SheetDocumento2 pagineAna Bruker Scion TQ Spec SheetmdananicNessuna valutazione finora
- Accurate Rapid Analysis of Alkali Contents in Portland CementDocumento4 pagineAccurate Rapid Analysis of Alkali Contents in Portland CementyinglvNessuna valutazione finora
- Valtam DataDocumento2 pagineValtam DataSantosh ZunjarNessuna valutazione finora
- 22.mohar Pal Meena GallDocumento11 pagine22.mohar Pal Meena GallVaidya NurNessuna valutazione finora
- Radiograph InterpretationDocumento12 pagineRadiograph InterpretationnithingrmeNessuna valutazione finora
- Lab Report Food Processing (Blanching)Documento5 pagineLab Report Food Processing (Blanching)siti irdinaNessuna valutazione finora
- Log InterpretationDocumento181 pagineLog InterpretationJorge Luis RamosNessuna valutazione finora
- Water Fueled CarDocumento12 pagineWater Fueled CarVenkata Harish Yadavalli100% (1)
- Flux 2331-Zx MsdsDocumento8 pagineFlux 2331-Zx Msdsibnu Groho Herry sampurnoNessuna valutazione finora
- Stabilization of Soil by Using Polypropylene FibersDocumento7 pagineStabilization of Soil by Using Polypropylene FibersIJRASETPublicationsNessuna valutazione finora
- Nerve Muscle PhysioDocumento30 pagineNerve Muscle Physiopatel_hanisha06Nessuna valutazione finora
- NYLON 6 (H20) : Perry Equipment CorporationDocumento2 pagineNYLON 6 (H20) : Perry Equipment CorporationZackNessuna valutazione finora
- Chemical Engineering Basics (INDIABIX)Documento1 paginaChemical Engineering Basics (INDIABIX)Kuo SarongNessuna valutazione finora
- H2 Quantum Physics - Part 1 Tutorial 2014 - StudentDocumento17 pagineH2 Quantum Physics - Part 1 Tutorial 2014 - StudentweinengxxNessuna valutazione finora
- (WWW - Entrance Exam - Net) Food TechnologyDocumento3 pagine(WWW - Entrance Exam - Net) Food Technologymohit gArhwalNessuna valutazione finora
- Barabrine SiDocumento2 pagineBarabrine SiRohan SahniNessuna valutazione finora
- Phase 1Documento32 paginePhase 1KamarajanNessuna valutazione finora
- Introduction To Structural DatabasesDocumento10 pagineIntroduction To Structural Databasessumit mahajanNessuna valutazione finora
- 7 - ThermochemistryDocumento31 pagine7 - ThermochemistryLeslie MasiyandimaNessuna valutazione finora
- A Study On Physico-Chemical Properties of Ground Water Quality of Various Locations of Kanpur CityDocumento3 pagineA Study On Physico-Chemical Properties of Ground Water Quality of Various Locations of Kanpur CityNeerja ShuklaNessuna valutazione finora
- LCD ScreenDocumento21 pagineLCD ScreenFagner Fernandes AraujoNessuna valutazione finora
- Jntuk SyllabusDocumento48 pagineJntuk SyllabusVenkateswarlu SivarathriNessuna valutazione finora
- Emr439r White Mica PDFDocumento41 pagineEmr439r White Mica PDFGabriel Erazo FierroNessuna valutazione finora
- Raghuvaran 2020 IOP Conf. Ser. Mater. Sci. Eng. 995 012040Documento9 pagineRaghuvaran 2020 IOP Conf. Ser. Mater. Sci. Eng. 995 012040Samuel ManoharNessuna valutazione finora
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4.5 su 5 stelle4.5/5 (6)
- Gut: the new and revised Sunday Times bestsellerDa EverandGut: the new and revised Sunday Times bestsellerValutazione: 4 su 5 stelle4/5 (393)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDa Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessValutazione: 4 su 5 stelle4/5 (33)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseDa EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseValutazione: 4.5 su 5 stelle4.5/5 (52)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDa EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedValutazione: 4 su 5 stelle4/5 (11)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDa EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldValutazione: 4 su 5 stelle4/5 (597)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDa EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNessuna valutazione finora
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDa EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsValutazione: 4.5 su 5 stelle4.5/5 (6)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDa EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesValutazione: 4.5 su 5 stelle4.5/5 (397)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDa EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceValutazione: 4.5 su 5 stelle4.5/5 (517)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDa EverandTales from Both Sides of the Brain: A Life in NeuroscienceValutazione: 3 su 5 stelle3/5 (18)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDa EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouValutazione: 4.5 su 5 stelle4.5/5 (62)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDa EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionValutazione: 4 su 5 stelle4/5 (812)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDa EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNessuna valutazione finora
- Who's in Charge?: Free Will and the Science of the BrainDa EverandWho's in Charge?: Free Will and the Science of the BrainValutazione: 4 su 5 stelle4/5 (65)
- Seven and a Half Lessons About the BrainDa EverandSeven and a Half Lessons About the BrainValutazione: 4 su 5 stelle4/5 (111)
- Good Without God: What a Billion Nonreligious People Do BelieveDa EverandGood Without God: What a Billion Nonreligious People Do BelieveValutazione: 4 su 5 stelle4/5 (66)
- Lymph & Longevity: The Untapped Secret to HealthDa EverandLymph & Longevity: The Untapped Secret to HealthValutazione: 4.5 su 5 stelle4.5/5 (13)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDa EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemValutazione: 4.5 su 5 stelle4.5/5 (116)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDa EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindValutazione: 4.5 su 5 stelle4.5/5 (93)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDa EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomValutazione: 4 su 5 stelle4/5 (216)
- Human: The Science Behind What Makes Your Brain UniqueDa EverandHuman: The Science Behind What Makes Your Brain UniqueValutazione: 3.5 su 5 stelle3.5/5 (38)
- Remnants of Ancient Life: The New Science of Old FossilsDa EverandRemnants of Ancient Life: The New Science of Old FossilsValutazione: 3 su 5 stelle3/5 (3)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDa EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondValutazione: 4 su 5 stelle4/5 (4)