Potrebbero piacerti anche
- Οξεοβασική ΙσορροπίαDocumento38 pagineΟξεοβασική ΙσορροπίαΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Enzyme Biotech Clinical ShortDocumento27 pagineEnzyme Biotech Clinical ShortΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- 02 Animations For 01Documento8 pagine02 Animations For 01Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Summary NB 2013Documento148 pagineSummary NB 2013Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Color Atlas of BiochemistryDocumento11 pagineColor Atlas of BiochemistryΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- 2 Purification 2013Documento61 pagine2 Purification 2013Μαρία-Άννα Κιρμπάκη100% (1)
- ΠεπτικόDocumento31 pagineΠεπτικόΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Allosteric Regulation - 2013Documento96 pagineAllosteric Regulation - 2013Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
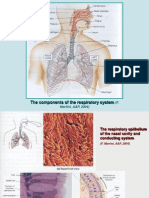
- The Components of The Respiratory System: (F. Martini, A&P, 2004)Documento23 pagineThe Components of The Respiratory System: (F. Martini, A&P, 2004)Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- ΜυϊκόDocumento40 pagineΜυϊκόΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Surfactant Composition and Function: Joanna Floros, PH.DDocumento21 pagineSurfactant Composition and Function: Joanna Floros, PH.DΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- Μεταγραφή και σύνθεση του RNADocumento70 pagineΜεταγραφή και σύνθεση του RNAΜαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- 1 Replication 2012Documento123 pagine1 Replication 2012Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- 1 Replication 2012Documento123 pagine1 Replication 2012Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- 3 RNA Maturation 2012Documento62 pagine3 RNA Maturation 2012Μαρία-Άννα ΚιρμπάκηNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5782)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (72)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Staar English I Short Answer Rubric-Connecting Selections 1pgDocumento1 paginaStaar English I Short Answer Rubric-Connecting Selections 1pgapi-268191635Nessuna valutazione finora
- Appendix 12.7.0 Fortinet Corporate ProfileDocumento8 pagineAppendix 12.7.0 Fortinet Corporate ProfileYan Naing ThwinNessuna valutazione finora
- Credit Seminar Final 08-05-2020-1Documento38 pagineCredit Seminar Final 08-05-2020-1Manoj Kumar LodhiNessuna valutazione finora
- Long-Run and Short-Run Concerns: Growth, Productivity, Unemployment, and in AtionDocumento34 pagineLong-Run and Short-Run Concerns: Growth, Productivity, Unemployment, and in AtionYunita AngelicaNessuna valutazione finora
- Parables of Jesus: The New Testament of The Holy Bible Contains Many Examples of The Parables of JesusDocumento3 pagineParables of Jesus: The New Testament of The Holy Bible Contains Many Examples of The Parables of JesuspaulbucudNessuna valutazione finora
- Survey Questionnaire NSTPDocumento7 pagineSurvey Questionnaire NSTPedwineiou83% (6)
- RAICES LawsuitDocumento25 pagineRAICES LawsuitJohn MoritzNessuna valutazione finora
- Dan Gilbert/ The Surprising Science of Happiness - TED TalkDocumento18 pagineDan Gilbert/ The Surprising Science of Happiness - TED TalkElenaNessuna valutazione finora
- BSF Witton ParkDocumento16 pagineBSF Witton ParkianbanksNessuna valutazione finora
- Appraisal Plan Template - SCAMPI B - V316Documento16 pagineAppraisal Plan Template - SCAMPI B - V316api-2626880150% (2)
- Лекция 7Documento76 pagineЛекция 7ramtin malekshahiNessuna valutazione finora
- Arduino Mega 2560 Microcontroller for Smart Car DesignDocumento2 pagineArduino Mega 2560 Microcontroller for Smart Car DesignAnil Kumar BNessuna valutazione finora
- Freepik BrandBook 07092023Documento84 pagineFreepik BrandBook 07092023Sai MoonNessuna valutazione finora
- Final Output in Intermediate Accounting Problems on Shareholders' EquityDocumento15 pagineFinal Output in Intermediate Accounting Problems on Shareholders' Equityennaira 06Nessuna valutazione finora
- CHEMICAL DISINFECTANTS AND THEIR MECHANISMS OF ACTIONDocumento29 pagineCHEMICAL DISINFECTANTS AND THEIR MECHANISMS OF ACTIONdadiboina gurusivaNessuna valutazione finora
- Partnership Agreement on Financing ProjectsDocumento12 paginePartnership Agreement on Financing ProjectsASSET GMBHNessuna valutazione finora
- SPA TO PROCESS (Loss OR CR)Documento1 paginaSPA TO PROCESS (Loss OR CR)majoy75% (4)
- Powerpoint PBL 2Documento13 paginePowerpoint PBL 2tariqNessuna valutazione finora
- Elementis Selector Chart AdditiveDocumento14 pagineElementis Selector Chart AdditiveEugene Pai100% (1)
- Caregiving 3Documento14 pagineCaregiving 3Bianca SandovalNessuna valutazione finora
- Term Paper: An Overview ToDocumento9 pagineTerm Paper: An Overview ToChing DialomaNessuna valutazione finora
- Manual Multilin 369Documento290 pagineManual Multilin 369Jesus Landaeta100% (1)
- Aakash-Bhora-Surja-Taara TabDocumento7 pagineAakash-Bhora-Surja-Taara TabKanai SinghaNessuna valutazione finora
- Slides 2Documento897 pagineSlides 2Samar QadirNessuna valutazione finora
- Progress Test 8 BDocumento2 pagineProgress Test 8 BEvaNessuna valutazione finora
- Inhibition in Speaking Ability of Fifth Semester Students in Class 2 of English Department at Tidar UniversityDocumento5 pagineInhibition in Speaking Ability of Fifth Semester Students in Class 2 of English Department at Tidar UniversityAzizi AziziNessuna valutazione finora
- Cranfield UniversityDocumento418 pagineCranfield UniversityElaf AhmedNessuna valutazione finora
- Bad Therapy: Matt Zoller SeitzDocumento3 pagineBad Therapy: Matt Zoller SeitzPatrik TeprakNessuna valutazione finora
- Bitwise Operations in CDocumento18 pagineBitwise Operations in CPriyam ChowdhuryNessuna valutazione finora
- The Committee To Scam New YorkDocumento31 pagineThe Committee To Scam New YorkpublicaccountabilityNessuna valutazione finora