Potrebbero piacerti anche
- Tsou & Fu 2007 - Octad Pollen Formation in Cymbopetalum (Annonaceae) - The Binding Mechanism PDFDocumento11 pagineTsou & Fu 2007 - Octad Pollen Formation in Cymbopetalum (Annonaceae) - The Binding Mechanism PDFMárcio BazanteNessuna valutazione finora
- Nematocitos PolypodiumDocumento11 pagineNematocitos PolypodiumKevin RucciNessuna valutazione finora
- Cito Esqueleto MeiosisDocumento10 pagineCito Esqueleto MeiosisYair Rodriguez SantiagoNessuna valutazione finora
- 16 Staehelin PDFDocumento12 pagine16 Staehelin PDFAmandaPutriNastitiNessuna valutazione finora
- Jackman and R. F. M. Van Steveninck : (Manuscript Received MayDocumento8 pagineJackman and R. F. M. Van Steveninck : (Manuscript Received MayMsMistiqueNessuna valutazione finora
- The Three-Dimensional Structure of The Cytostome-CytopharynxDocumento11 pagineThe Three-Dimensional Structure of The Cytostome-CytopharynxUNIG Aluana SantanaNessuna valutazione finora
- Crop Physiology Exercise-2Documento7 pagineCrop Physiology Exercise-2Ravikant MishraNessuna valutazione finora
- New Phytologist - 2008 - SKENE - Cluster Root Development in Grevillea Robusta Proteaceae I Xylem Pericycle CortexDocumento8 pagineNew Phytologist - 2008 - SKENE - Cluster Root Development in Grevillea Robusta Proteaceae I Xylem Pericycle Cortexyyangg.0802Nessuna valutazione finora
- Southworth Et Al 2001-Male GametogenesisDocumento16 pagineSouthworth Et Al 2001-Male GametogenesisAde MulyanaNessuna valutazione finora
- 2229 FullDocumento7 pagine2229 FullRifqi Thirafi RamadhanNessuna valutazione finora
- Bacteria TermitasDocumento8 pagineBacteria TermitasRashaverak RomNessuna valutazione finora
- Research in ProtozoologyDa EverandResearch in ProtozoologyTze-Tuan ChenNessuna valutazione finora
- Aplanogamous Sexual Reproduction in Carteria Eugametos Volvocales ChlorophytaDocumento6 pagineAplanogamous Sexual Reproduction in Carteria Eugametos Volvocales ChlorophytaSirNessuna valutazione finora
- 04 Prokaryotic Algae ProcholopytesDocumento17 pagine04 Prokaryotic Algae Procholopytesanne ropNessuna valutazione finora
- TMP A6 AFDocumento9 pagineTMP A6 AFFrontiersNessuna valutazione finora
- Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on EarthDa EverandKingdoms and Domains: An Illustrated Guide to the Phyla of Life on EarthValutazione: 4.5 su 5 stelle4.5/5 (4)
- Developmental Anatomy of Adventitious Shoot Formation On Snapdragon (Antirrhinum Majus L.) Hypocotyls in VitroDocumento5 pagineDevelopmental Anatomy of Adventitious Shoot Formation On Snapdragon (Antirrhinum Majus L.) Hypocotyls in VitroFrancisco Moctezuma EsquivelNessuna valutazione finora
- Caveolae AdipocytesDocumento10 pagineCaveolae AdipocytesReza AzghadiNessuna valutazione finora
- Embriologi T. PaniculatumDocumento24 pagineEmbriologi T. Paniculatumfanny dominggaNessuna valutazione finora
- Aamir Nawaz KhanDocumento20 pagineAamir Nawaz KhanBslssbxjxnbbxNessuna valutazione finora
- Area Opaca (Figures 1, 2) - The Latter Region Contributes Only To ExtraembryonicDocumento13 pagineArea Opaca (Figures 1, 2) - The Latter Region Contributes Only To ExtraembryonicgiuseppegnrNessuna valutazione finora
- On The Origins of Mitosing Cells - 1967 PDFDocumento56 pagineOn The Origins of Mitosing Cells - 1967 PDFMartín FuentesNessuna valutazione finora
- 1 s2.0 0022519367900793 Main 1 1 25Documento25 pagine1 s2.0 0022519367900793 Main 1 1 25Dan Gerson Apaza AracayoNessuna valutazione finora
- A Comparative Cytological Analysis of Fu20160623 13956 q2w8t5 With Cover PageDocumento15 pagineA Comparative Cytological Analysis of Fu20160623 13956 q2w8t5 With Cover PageCristián MaximilianoNessuna valutazione finora
- Entomoneis Vertebralis Sp. Nov. (Bacillariophyceae) A New Species From Hypersaline EnvironmentsDocumento12 pagineEntomoneis Vertebralis Sp. Nov. (Bacillariophyceae) A New Species From Hypersaline EnvironmentsÝ NhiNessuna valutazione finora
- Sagan, L. On The Origin of Mitosing Cells. J. Theoret. Biol. (1967) 14, 225-274Documento56 pagineSagan, L. On The Origin of Mitosing Cells. J. Theoret. Biol. (1967) 14, 225-274Jéssica CristinaNessuna valutazione finora
- Sdarticle 32Documento8 pagineSdarticle 32api-19973331Nessuna valutazione finora
- Raviteja1 PDFDocumento6 pagineRaviteja1 PDFMekala LakshmanNessuna valutazione finora
- Mind Blindness and The Brain in AutismDocumento4 pagineMind Blindness and The Brain in AutismRenata Lima de SantanaNessuna valutazione finora
- 1 ForaminiferaDocumento40 pagine1 ForaminiferaFrancisco JavierNessuna valutazione finora
- PhaeophyceaeDocumento4 paginePhaeophyceaeAdi WeerasingheNessuna valutazione finora
- ChloroplastDocumento2 pagineChloroplastNoraini RahimNessuna valutazione finora
- Observing Microscopic Phases of Lichen Life Cycles On Transparent Substrata Placed in SituDocumento10 pagineObserving Microscopic Phases of Lichen Life Cycles On Transparent Substrata Placed in SituCinthia Edith Huamani GamonalNessuna valutazione finora
- Cellulose - The Structure Slowly UnravelsDocumento35 pagineCellulose - The Structure Slowly UnravelsAditya Shrivastava100% (1)
- Merizocotyle Australensis Himantura FaiDocumento12 pagineMerizocotyle Australensis Himantura FaiAlexander MoralesNessuna valutazione finora
- CAPE PhotosynthesisDocumento20 pagineCAPE PhotosynthesisIsheba Warren100% (24)
- Fine Structure and Morphogenic Movements in The Gastrula of The Treefrog, Hyla RegillaDocumento24 pagineFine Structure and Morphogenic Movements in The Gastrula of The Treefrog, Hyla Regillaimanuel kuneNessuna valutazione finora
- Bannister Et Al 1986 Merozoite Membrane ThicknesDocumento10 pagineBannister Et Al 1986 Merozoite Membrane ThicknesArunaditya Deshmukh (Adi)Nessuna valutazione finora
- OCES3003 Lab 7 Zooplanktkon GrazingDocumento4 pagineOCES3003 Lab 7 Zooplanktkon GrazingNokyin ChanNessuna valutazione finora
- Vakuola Kontraktil PDFDocumento17 pagineVakuola Kontraktil PDFShin Oriflame MalangNessuna valutazione finora
- Kalinina Protistology 12-1Documento9 pagineKalinina Protistology 12-1jesus javier hoyos acuñaNessuna valutazione finora
- Cytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDocumento11 pagineCytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDario DiosNessuna valutazione finora
- Scan Tugas Rumbuhan AirDocumento4 pagineScan Tugas Rumbuhan AirRyan TamherNessuna valutazione finora
- Porifera: Sally P. Leys and Nathan FarrarDocumento9 paginePorifera: Sally P. Leys and Nathan FarrarFaradilla Aqidatul IzzahNessuna valutazione finora
- CHP 2 Structure and Function of CellsDocumento29 pagineCHP 2 Structure and Function of CellsFathima NusrathNessuna valutazione finora
- Montagnac 2008Documento8 pagineMontagnac 2008JUSTINE CHLOE DEL ROSARIONessuna valutazione finora
- Jbacter00584 0499Documento14 pagineJbacter00584 0499YO :)Nessuna valutazione finora
- S Asamoto 1992Documento5 pagineS Asamoto 1992DANIEL ARTEAGA MENDOZANessuna valutazione finora
- (Late Triassic) of On The Origin of The Angiosperm Carpel and StamenDocumento36 pagine(Late Triassic) of On The Origin of The Angiosperm Carpel and StamenMolinaArzabeCarlos100% (1)
- Microfossils of Pulau Salakan, SabahDocumento18 pagineMicrofossils of Pulau Salakan, SabahAhmad LadoNessuna valutazione finora
- The Distribution of TPX2 in Dividing Leaf Cells of The Fern Asplenium NidusDocumento7 pagineThe Distribution of TPX2 in Dividing Leaf Cells of The Fern Asplenium Nidusayyub ramadhanNessuna valutazione finora
- Cell Evolution and Earth History 2006Documento38 pagineCell Evolution and Earth History 2006Iván LazcanoNessuna valutazione finora
- Pollen Tube in PetuniaDocumento20 paginePollen Tube in PetuniaevayrbtpNessuna valutazione finora
- An Electron Microscope Study of Thin Sections of Human Skin LPADocumento21 pagineAn Electron Microscope Study of Thin Sections of Human Skin LPALucas PolizzeliNessuna valutazione finora
- Cytoplast Concept in Dividing Cells Pickett-Heaps AmJBota1999Documento20 pagineCytoplast Concept in Dividing Cells Pickett-Heaps AmJBota1999AJMRNessuna valutazione finora
- 1 ForaminiferaDocumento51 pagine1 ForaminiferaManoj KitchappanNessuna valutazione finora
- General Characteristics of CyanophyceaeDocumento27 pagineGeneral Characteristics of Cyanophyceaepreethisutha29Nessuna valutazione finora
- Cross Talk Details and RoutingDocumento29 pagineCross Talk Details and RoutingRohith RajNessuna valutazione finora
- Data Bulletin Group Motor Installations:: Understanding National Electrical Code (NEC) 430.53 RequirementsDocumento8 pagineData Bulletin Group Motor Installations:: Understanding National Electrical Code (NEC) 430.53 RequirementsshoaibNessuna valutazione finora
- Esteem 1999 2000 1.3L 1.6LDocumento45 pagineEsteem 1999 2000 1.3L 1.6LArnold Hernández CarvajalNessuna valutazione finora
- Material Specification - 077154C-000-JSS-1700-009 - DDocumento13 pagineMaterial Specification - 077154C-000-JSS-1700-009 - DStructures ProductionNessuna valutazione finora
- LET General Math ReviewerDocumento7 pagineLET General Math ReviewerMarco Rhonel Eusebio100% (1)
- The Broadband ForumDocumento21 pagineThe Broadband ForumAnouar AleyaNessuna valutazione finora
- Innerwear Industry Pitch PresentationDocumento19 pagineInnerwear Industry Pitch PresentationRupeshKumarNessuna valutazione finora
- Prestige Institute of Management & Research: Guided By:-Submitted By: - Prof. Arpit Loya Sumeet RattanDocumento21 paginePrestige Institute of Management & Research: Guided By:-Submitted By: - Prof. Arpit Loya Sumeet RattanSumeet700005Nessuna valutazione finora
- Syllabus Unit Iv Unit Commitment and Economic DispatchDocumento23 pagineSyllabus Unit Iv Unit Commitment and Economic DispatchBALAKRISHNANNessuna valutazione finora
- Niir Integrated Organic Farming Handbook PDFDocumento13 pagineNiir Integrated Organic Farming Handbook PDFNatalieNessuna valutazione finora
- Plastics and Polymer EngineeringDocumento4 paginePlastics and Polymer Engineeringsuranjana26Nessuna valutazione finora
- Course Syllabus: Course Code Course Title ECTS CreditsDocumento3 pagineCourse Syllabus: Course Code Course Title ECTS CreditsHanaa HamadallahNessuna valutazione finora
- Bulacan Agricultural State College: Lesson Plan in Science 4 Life Cycle of Humans, Animals and PlantsDocumento6 pagineBulacan Agricultural State College: Lesson Plan in Science 4 Life Cycle of Humans, Animals and PlantsHarmonica PellazarNessuna valutazione finora
- CheckList For Checking of Drawings-R1Documento4 pagineCheckList For Checking of Drawings-R1jatin kalraNessuna valutazione finora
- Cynosure Starlux 500 Palomar Technical Service ManualDocumento47 pagineCynosure Starlux 500 Palomar Technical Service ManualJF SilvaNessuna valutazione finora
- NF en Iso 5167-6-2019Documento22 pagineNF en Iso 5167-6-2019Rem FgtNessuna valutazione finora
- Arduino Oscilloscope ProjectDocumento12 pagineArduino Oscilloscope ProjectSathya Narayan100% (1)
- 5G Transport Slice Control in End-To-End 5G NetworksDocumento19 pagine5G Transport Slice Control in End-To-End 5G NetworksmorganNessuna valutazione finora
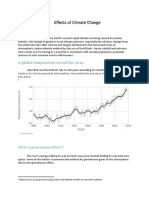
- Effects of Climate ChangeDocumento3 pagineEffects of Climate Changejiofjij100% (1)
- Advanta V12 Data SheetDocumento2 pagineAdvanta V12 Data SheetJuliana MiyagiNessuna valutazione finora
- Middle Range Theory Ellen D. Schulzt: Modeling and Role Modeling Katharine Kolcaba: Comfort TheoryDocumento22 pagineMiddle Range Theory Ellen D. Schulzt: Modeling and Role Modeling Katharine Kolcaba: Comfort TheoryMerlinNessuna valutazione finora
- Digging Deeper: Can Hot Air Provide Sustainable Source of Electricity?Documento2 pagineDigging Deeper: Can Hot Air Provide Sustainable Source of Electricity?Рустам ХаджаевNessuna valutazione finora
- Lesson 1 Chapter 9 ErosiondepositionDocumento1 paginaLesson 1 Chapter 9 Erosiondepositionapi-249320969Nessuna valutazione finora
- Impact of Retrofitting Existing Combined Heat and Power Plant With Polygeneration of Biomethane PDFDocumento16 pagineImpact of Retrofitting Existing Combined Heat and Power Plant With Polygeneration of Biomethane PDFAwais Salman0% (1)
- EXP4 The Diels Alder ReactionsDocumento3 pagineEXP4 The Diels Alder ReactionsLaura GuidoNessuna valutazione finora
- Electrowetting - Wikipedia, The Free EncyclopediaDocumento5 pagineElectrowetting - Wikipedia, The Free EncyclopediaDwane AlmeidaNessuna valutazione finora
- 12 Key Principles On Bhava AnalysisDocumento28 pagine12 Key Principles On Bhava AnalysisDhineshNessuna valutazione finora
- The FOA Reference For Fiber Optics - Fiber Optic TestingDocumento19 pagineThe FOA Reference For Fiber Optics - Fiber Optic TestingvsalaiselvamNessuna valutazione finora
- Significance of GodboleDocumento5 pagineSignificance of GodbolehickeyvNessuna valutazione finora
- 07ercoskun 05 01 PDFDocumento23 pagine07ercoskun 05 01 PDFjagmadridNessuna valutazione finora