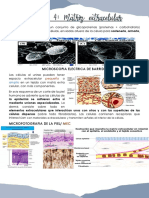
Potrebbero piacerti anche
- Tipos de uniones en la adherencia celularDocumento14 pagineTipos de uniones en la adherencia celularAndy Paúl García OrbeNessuna valutazione finora
- Moleculas de AdhesionDocumento10 pagineMoleculas de AdhesionTefita XivitaNessuna valutazione finora
- Molécula de Adhesión CelularDocumento20 pagineMolécula de Adhesión CelularSkipy RosalesNessuna valutazione finora
- MAC: Moléculas de adhesión celularDocumento20 pagineMAC: Moléculas de adhesión celularMarce De La CruzNessuna valutazione finora
- Moleculas de Adhesion y PielDocumento13 pagineMoleculas de Adhesion y PielJesus Bautisto Huichi JaraNessuna valutazione finora
- Moleculas de Adhecion CelularDocumento3 pagineMoleculas de Adhecion CelularAngel Daniel Ocampo VazquezNessuna valutazione finora
- Moléculas de Adhesión CelularDocumento7 pagineMoléculas de Adhesión CelularAbel Gallegos AguilarNessuna valutazione finora
- Molécula de Adhesión CelularDocumento4 pagineMolécula de Adhesión CelularcristiaanchNessuna valutazione finora
- Biología Celular y Molecular - SeminarioDocumento13 pagineBiología Celular y Molecular - SeminarioTEFANessuna valutazione finora
- IntegrinasDocumento8 pagineIntegrinasAlejandra Cueva FloresNessuna valutazione finora
- MetabolismoDocumento17 pagineMetabolismoFernando J. Llanganate DNessuna valutazione finora
- Moleculas de Adhecion CelularDocumento4 pagineMoleculas de Adhecion CelularAnders Kevin Lagos ValenzuelaNessuna valutazione finora
- Moléculas de Relación CelularDocumento15 pagineMoléculas de Relación CelularAlejandro SuaresNessuna valutazione finora
- LectinasDocumento10 pagineLectinascarlosventosaNessuna valutazione finora
- Chaperon AsDocumento35 pagineChaperon Aslord1992Nessuna valutazione finora
- Notas Sobre MetabolismoDocumento23 pagineNotas Sobre MetabolismoPerla PereiraNessuna valutazione finora
- TP 4 RespuestasDocumento13 pagineTP 4 RespuestasLuciana E RomeroNessuna valutazione finora
- Metabolismo 40Documento13 pagineMetabolismo 40Carmen Carolina Ruiz GarzaNessuna valutazione finora
- 2-Proteinas Parte 1Documento7 pagine2-Proteinas Parte 1anonimo.aaa1231998Nessuna valutazione finora
- Lectinas Vegetales ArticuloDocumento7 pagineLectinas Vegetales Articuloarcadia_hbNessuna valutazione finora
- U4 PP 88 MetabolismoDocumento19 pagineU4 PP 88 MetabolismoCarlos SalernoNessuna valutazione finora
- Introducción Al Metabolismo PDFDocumento10 pagineIntroducción Al Metabolismo PDFJhonCastro182Nessuna valutazione finora
- Endotelio Vascular y La Circulación de Los LeucocitosDocumento20 pagineEndotelio Vascular y La Circulación de Los LeucocitosEveGarCoNessuna valutazione finora
- Rutas CatabolicasDocumento13 pagineRutas CatabolicasCarlos david Altares AltamiraNessuna valutazione finora
- Interacción Entre Las Células y Su EntornoDocumento8 pagineInteracción Entre Las Células y Su EntornoAnita EspinosaNessuna valutazione finora
- Resumen Del Primer Parcial de Tecno 224Documento30 pagineResumen Del Primer Parcial de Tecno 224Dayana PizarroNessuna valutazione finora
- Adherencia CelularDocumento35 pagineAdherencia CelularRosmeri RomeroNessuna valutazione finora
- Informe 8 Bio Sem 2020Documento10 pagineInforme 8 Bio Sem 2020anton cabreraNessuna valutazione finora
- METABOLISMODocumento20 pagineMETABOLISMOBolonia Fiambres y QuesosNessuna valutazione finora
- Recirculacion de LinfocitosDocumento6 pagineRecirculacion de LinfocitosjankitaNessuna valutazione finora
- 1QM1 22-1 Juarez Paola Avance de CuestionariosDocumento10 pagine1QM1 22-1 Juarez Paola Avance de CuestionariosPaola JuarezNessuna valutazione finora
- Metabolismo - Wikipedia, La Enciclopedia LibreDocumento142 pagineMetabolismo - Wikipedia, La Enciclopedia LibreDalwin GilNessuna valutazione finora
- S5-Matriz Celular ExtracelularDocumento18 pagineS5-Matriz Celular ExtracelularJhony Farith Farfán ReáteguiNessuna valutazione finora
- X El MicroambienteDocumento6 pagineX El MicroambienteDiana Karen Butanda PuenteNessuna valutazione finora
- Interacciones de Las Células Con Los Materiales ExtracelularesDocumento7 pagineInteracciones de Las Células Con Los Materiales ExtracelularesFlor CarreraNessuna valutazione finora
- Matriz ExtracelularDocumento36 pagineMatriz ExtracelularArmando BravoNessuna valutazione finora
- Capítulo 26 Descubrimiento y Clasificación de Las Proteínas de Unión A GlicanoDocumento14 pagineCapítulo 26 Descubrimiento y Clasificación de Las Proteínas de Unión A GlicanoLizeth Jocelyn Serna VillalobosNessuna valutazione finora
- Bio Sem InformeDocumento15 pagineBio Sem InformeXiomara Janne Linares SaldañaNessuna valutazione finora
- Transporte CelularDocumento24 pagineTransporte CelularIvonne Andrea Donoso ToledoNessuna valutazione finora
- Uso de Lectinas en IHDocumento5 pagineUso de Lectinas en IHVivi SaldiviaNessuna valutazione finora
- Clase 6 InmunohematologiaDocumento49 pagineClase 6 InmunohematologiareneNessuna valutazione finora
- Guía de taller sobre adhesión celularDocumento12 pagineGuía de taller sobre adhesión celularKAREN MONSERRAT FRANCO ACHARNessuna valutazione finora
- Membrana proteínasDocumento3 pagineMembrana proteínasABIGAIL ESPINOZANessuna valutazione finora
- METABOLISMODocumento86 pagineMETABOLISMOAlejandro ArreazaNessuna valutazione finora
- Catabolismo y AnabolismoDocumento4 pagineCatabolismo y AnabolismoKevin PachecoNessuna valutazione finora
- Moleculas de Adhesion Celular 1223605666724403 9Documento54 pagineMoleculas de Adhesion Celular 1223605666724403 9Karen CabanaNessuna valutazione finora
- Biologia Selectividad Examen 10 Resuelto Castilla La Mancha WWW - Siglo21x.blogspotDocumento7 pagineBiologia Selectividad Examen 10 Resuelto Castilla La Mancha WWW - Siglo21x.blogspotmonografiasx0% (1)
- 1QM1 22-1 Pérez Espinosa Luz Tonantzin Cuestionario Cuarta UnidadDocumento10 pagine1QM1 22-1 Pérez Espinosa Luz Tonantzin Cuestionario Cuarta UnidadLuz Tonantzin Pérez EspjnosaNessuna valutazione finora
- Seminario de Citoquinas-InmunologiaDocumento18 pagineSeminario de Citoquinas-InmunologiaMiguel MontillaNessuna valutazione finora
- Biologia Celular Tema 3.2 y 3.3Documento68 pagineBiologia Celular Tema 3.2 y 3.3christian GarciaNessuna valutazione finora
- Migración de Los Leucocitos A Los TejidosDocumento8 pagineMigración de Los Leucocitos A Los TejidosDiana Figueroa67% (3)
- Biologia Teoria Pregunta 6 y 7 Taller 2Documento7 pagineBiologia Teoria Pregunta 6 y 7 Taller 2estefaniaNessuna valutazione finora
- Integrinas y SelectinasDocumento3 pagineIntegrinas y SelectinasDiego Azamat100% (1)
- Exposicion de BioDocumento7 pagineExposicion de BioOsneider Peña CuetoNessuna valutazione finora
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesDa EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesValutazione: 5 su 5 stelle5/5 (1)
- 36 Profilaxis de AccidenteDocumento6 pagine36 Profilaxis de AccidentecopepinaNessuna valutazione finora
- Cocina para Bebes Con BarneyDocumento97 pagineCocina para Bebes Con BarneycopepinaNessuna valutazione finora
- Estimulación para Niños de 2 y 3 AñosDocumento13 pagineEstimulación para Niños de 2 y 3 Añosmatildedelatorre_muriedasNessuna valutazione finora
- Cocina para Bebes Con BarneyDocumento97 pagineCocina para Bebes Con BarneycopepinaNessuna valutazione finora
- MBE ElectronicaDocumento13 pagineMBE ElectronicacopepinaNessuna valutazione finora
- Cascada Del Complemento InmunologicoDocumento16 pagineCascada Del Complemento Inmunologicomiguel angel100% (30)
- MBE Hacer La PreguntaDocumento3 pagineMBE Hacer La PreguntacopepinaNessuna valutazione finora
- Mbe Niveles EvdenciaDocumento2 pagineMbe Niveles EvdenciacopepinaNessuna valutazione finora
- Normas técnicas nacionales sobre residuos biopatológicosDocumento8 pagineNormas técnicas nacionales sobre residuos biopatológicoscopepinaNessuna valutazione finora
- 404 PH Metros y Otros Instrumentos de Medida ElectroquimicaDocumento4 pagine404 PH Metros y Otros Instrumentos de Medida ElectroquimicacopepinaNessuna valutazione finora
- Leishmaniasis DX PDFDocumento45 pagineLeishmaniasis DX PDFmeds1313Nessuna valutazione finora
- AvidezDocumento5 pagineAvidezcopepinaNessuna valutazione finora
- Actividades para Desarrollar La Ejecución Verbal. Centro Concertado de Educación Especial Ángel Riviére.Documento23 pagineActividades para Desarrollar La Ejecución Verbal. Centro Concertado de Educación Especial Ángel Riviére.bichitavetNessuna valutazione finora
- Qca. Biologica 5.27 El Metabolismo IntermediarioDocumento4 pagineQca. Biologica 5.27 El Metabolismo IntermediariocopepinaNessuna valutazione finora
- Libro Parejas TormentosasDocumento99 pagineLibro Parejas TormentosasKaren ArteagaNessuna valutazione finora
- 000 Pruebas BioquimicasDocumento3 pagine000 Pruebas Bioquimicascopepina100% (1)
- Alta Recetas EnsurepolvoDocumento4 pagineAlta Recetas EnsurepolvoMariella V. IshiharaNessuna valutazione finora
- Infeccion Por Toxoplasma Gondii Durante El EmbarazoDocumento4 pagineInfeccion Por Toxoplasma Gondii Durante El EmbarazocopepinaNessuna valutazione finora
- 400 PotenciometriaDocumento8 pagine400 PotenciometriacopepinaNessuna valutazione finora
- 12 Activacion de Celulas NKDocumento15 pagine12 Activacion de Celulas NKcopepinaNessuna valutazione finora
- GRUPOS SANGUINEO ABO Y RHDocumento12 pagineGRUPOS SANGUINEO ABO Y RHJesús LozanoNessuna valutazione finora
- Anatomía del movimiento: planos, ejes y palancasDocumento7 pagineAnatomía del movimiento: planos, ejes y palancasAntar Jose Gòmez CardeñaNessuna valutazione finora
- Cultivo Primario de Embrión de Pollo.Documento4 pagineCultivo Primario de Embrión de Pollo.copepinaNessuna valutazione finora
- 3.15 ReticulocitosDocumento4 pagine3.15 ReticulocitoscopepinaNessuna valutazione finora
- Mecanismos de Evasio de La Respuesta InmuneDocumento10 pagineMecanismos de Evasio de La Respuesta InmuneMariano Hernandez PimentelNessuna valutazione finora
- 3.11 Recuento de Leucocitos en CamaraDocumento4 pagine3.11 Recuento de Leucocitos en CamaracopepinaNessuna valutazione finora
- 002 Retrovirus EditorialDocumento3 pagine002 Retrovirus EditorialcopepinaNessuna valutazione finora
- Efecto CitopaticoDocumento7 pagineEfecto CitopaticoJonathan Manuel Jimenez LunaNessuna valutazione finora
- 003 Virus y Las Enfermedades - CampusDocumento14 pagine003 Virus y Las Enfermedades - CampuscopepinaNessuna valutazione finora
- Celula VegetalDocumento41 pagineCelula VegetalLeonor Ramos100% (1)
- P1 Morfofisiología IDocumento13 pagineP1 Morfofisiología IIsbelis Flores100% (3)
- Requerimientos Nutricionales Del Caballo NRC 6ta EdDocumento1 paginaRequerimientos Nutricionales Del Caballo NRC 6ta EdErnest HemilNessuna valutazione finora
- Tiroides, Maestria 2013Documento131 pagineTiroides, Maestria 2013Carlos Buitrago100% (1)
- Presencia de Alcaloides en Plantas MedicinalesDocumento3 paginePresencia de Alcaloides en Plantas MedicinalesMaribel PerezNessuna valutazione finora
- LeucotrienosDocumento9 pagineLeucotrienosDiana JuarezNessuna valutazione finora
- PCCNS para TutorDocumento465 paginePCCNS para TutorKristell DerasNessuna valutazione finora
- Bases químicas de la vida: guía de estudio para práctica de biología generalDocumento3 pagineBases químicas de la vida: guía de estudio para práctica de biología generalEylin SalgueroNessuna valutazione finora
- Acción de la bromelina e identificación de aminoácidos en gelatinaDocumento8 pagineAcción de la bromelina e identificación de aminoácidos en gelatinaFernandoNessuna valutazione finora
- Histología: Estudio visual de tejidos a nivel microscópicoDocumento134 pagineHistología: Estudio visual de tejidos a nivel microscópicoDani Morales DellatorreNessuna valutazione finora
- Fotosíntesis y respiraciónDocumento4 pagineFotosíntesis y respiraciónKely NúñezNessuna valutazione finora
- Infografía Respuesta Al TraumaDocumento1 paginaInfografía Respuesta Al TraumaEl Cesar 55Nessuna valutazione finora
- Estudio del bombeo de protones por levadurasDocumento5 pagineEstudio del bombeo de protones por levadurasAlondra FortunatNessuna valutazione finora
- Metabolismo y EnergíaDocumento42 pagineMetabolismo y EnergíaJesús Rondón RíosNessuna valutazione finora
- GluconeogénesisDocumento16 pagineGluconeogénesisGiovanni GonzalezNessuna valutazione finora
- Evaluación de CélulaDocumento2 pagineEvaluación de CélulaCarlos LuboNessuna valutazione finora
- Guia de Laboratorio EnzimasDocumento3 pagineGuia de Laboratorio EnzimasNalda Pillco ChicllaNessuna valutazione finora
- Bioquimica Cuarto FolletoDocumento5 pagineBioquimica Cuarto FolletofredyNessuna valutazione finora
- Guia 3Documento22 pagineGuia 3Andrea NaviaNessuna valutazione finora
- Pared CelularDocumento5 paginePared CelularColumba LopezNessuna valutazione finora
- Regulación miRNA's células SchwannDocumento4 pagineRegulación miRNA's células SchwannMaría José García infanteNessuna valutazione finora
- Resumen Digestivo AnimalDocumento5 pagineResumen Digestivo Animalstefannyges100% (1)
- Célula Vegetal y Sus PartesDocumento8 pagineCélula Vegetal y Sus PartesELIA ROSASNessuna valutazione finora
- Bitacora de La Practica 6Documento3 pagineBitacora de La Practica 6Nathan Ulises Reyes JimenezNessuna valutazione finora
- Introduccion A La BiologiaDocumento11 pagineIntroduccion A La BiologiaAlanna De LeónNessuna valutazione finora
- Reconocimiento ProteinasDocumento19 pagineReconocimiento ProteinasNerlyn Santa FlorezNessuna valutazione finora
- KKKKKK Aa L1.1Documento11 pagineKKKKKK Aa L1.1PeaceSoldierNessuna valutazione finora
- ANTITUBERCULOSOSDocumento1 paginaANTITUBERCULOSOSdannyNessuna valutazione finora
- Taller Ciclo Celular B. GeneralDocumento4 pagineTaller Ciclo Celular B. GeneralDavid Vergara MorenoNessuna valutazione finora
- Test Examen Completo BQDocumento114 pagineTest Examen Completo BQAlberto BalbinNessuna valutazione finora