Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Activated Carbon For Purification of AlcoholDocumento28 pagineActivated Carbon For Purification of AlcoholcmdcscribdNessuna valutazione finora
- Restore Well Productivity with M-I PC Scale DissolversDocumento57 pagineRestore Well Productivity with M-I PC Scale Dissolversjps21100% (2)
- Charcoal RefDocumento7 pagineCharcoal RefAmit KumarNessuna valutazione finora
- Vitamin Requirement As Per NRCDocumento9 pagineVitamin Requirement As Per NRCAmit KumarNessuna valutazione finora
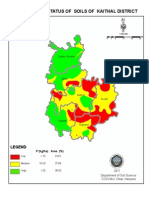
- Phosphorus Status of Soils of Kaithal District: LegendDocumento1 paginaPhosphorus Status of Soils of Kaithal District: LegendAmit KumarNessuna valutazione finora
- Copper RoleDocumento8 pagineCopper RoleAmit KumarNessuna valutazione finora
- Mecovet Dose ReferenceDocumento4 pagineMecovet Dose ReferenceAmit KumarNessuna valutazione finora
- Presentation NutritionDocumento82 paginePresentation NutritionAmit KumarNessuna valutazione finora
- Erythrocyte replacement and Methylcobalamin deficiency anemiaDocumento1 paginaErythrocyte replacement and Methylcobalamin deficiency anemiaAmit KumarNessuna valutazione finora
- Drugs ListDocumento195 pagineDrugs ListAmit Kumar100% (3)
- Fillers For Paints - GysauDocumento319 pagineFillers For Paints - GysaukiddprofessorNessuna valutazione finora
- Vitamin D and CalciumDocumento33 pagineVitamin D and CalciumAkhmadRoziNessuna valutazione finora
- MSDS Calsium OxideDocumento6 pagineMSDS Calsium OxidePipit RatnasariNessuna valutazione finora
- Gayatri Fertiplants International Pvt. LTDDocumento42 pagineGayatri Fertiplants International Pvt. LTDPassmore DubeNessuna valutazione finora
- TERHAD document physical properties states matter identification reactionsDocumento2 pagineTERHAD document physical properties states matter identification reactionsjanizanNessuna valutazione finora
- National Institute of Technology Raipur: Bio-ConcreteDocumento19 pagineNational Institute of Technology Raipur: Bio-ConcreteRahul DhakerNessuna valutazione finora
- Piochem Catalogue PDFDocumento98 paginePiochem Catalogue PDFمحمد التابعيNessuna valutazione finora
- ERW Hook CrackDocumento9 pagineERW Hook CrackElias KapaNessuna valutazione finora
- Calcium Silicate Insulation BrochureDocumento52 pagineCalcium Silicate Insulation BrochuremukeshsinghtomarNessuna valutazione finora
- A Novel Process For Extracting Lithium From Lepidolite 2012 HydrometallurgyDocumento6 pagineA Novel Process For Extracting Lithium From Lepidolite 2012 HydrometallurgyRose Olivares PinoNessuna valutazione finora
- VA GroupDocumento3 pagineVA GroupvipulNessuna valutazione finora
- DG of Chemical Reaction and Its EffectDocumento23 pagineDG of Chemical Reaction and Its EffectAnshuman MishraNessuna valutazione finora
- DD Ebook Deco Treatment PDFDocumento40 pagineDD Ebook Deco Treatment PDFFare NienteNessuna valutazione finora
- Water Impact Index: Evaluating the Impact of Human Activity on Water ResourcesDocumento33 pagineWater Impact Index: Evaluating the Impact of Human Activity on Water ResourcesSunny OoiNessuna valutazione finora
- Chemistry and Uses of Dolomite MineralDocumento11 pagineChemistry and Uses of Dolomite MineralMeitri Wulandari Kohar100% (1)
- US20150086452A1Documento11 pagineUS20150086452A1Alfianto AlfiantoNessuna valutazione finora
- As Level Chemistry: Topic 2 - Amount of Substance Assessed HomeworkDocumento15 pagineAs Level Chemistry: Topic 2 - Amount of Substance Assessed Homeworkkarima akterNessuna valutazione finora
- Everything You Need to Know About Portland CementDocumento23 pagineEverything You Need to Know About Portland CementSIDRA NAZEER SAIFNessuna valutazione finora
- 2009 UNIT 1 Paper 1Documento5 pagine2009 UNIT 1 Paper 1cilacax404Nessuna valutazione finora
- Fapas Water and Environmental (Leap) Programme 2016Documento30 pagineFapas Water and Environmental (Leap) Programme 2016linamendozaNessuna valutazione finora
- Valency: Combining Power of AtomsDocumento56 pagineValency: Combining Power of AtomsDaniel PalmerNessuna valutazione finora
- Quick Lime Powder ReportDocumento6 pagineQuick Lime Powder Reportnasir ahmedNessuna valutazione finora
- Fuji Dri-Chem Slide Ca-Piii: Date of Issue: 1/may/2012Documento1 paginaFuji Dri-Chem Slide Ca-Piii: Date of Issue: 1/may/2012susey tepaNessuna valutazione finora
- Periodic-Table-Groups B v6 Anm s1Documento31 paginePeriodic-Table-Groups B v6 Anm s1Sophi VijayNessuna valutazione finora
- Soil Stabilization With Rice Husk AshDocumento20 pagineSoil Stabilization With Rice Husk AshOkolo ValentineNessuna valutazione finora
- PE 1 Grade 12Documento8 paginePE 1 Grade 12Jandel CabildoNessuna valutazione finora
- Cannabis Nutrient Deficiency ChartsDocumento4 pagineCannabis Nutrient Deficiency Chartsa6587055100% (1)
- Solved General Science & Ability CSS Paper 2014 - MCQs With ExplanationDocumento16 pagineSolved General Science & Ability CSS Paper 2014 - MCQs With ExplanationUsman KhanNessuna valutazione finora
- Chapter 3Documento76 pagineChapter 3Eumar FabruadaNessuna valutazione finora