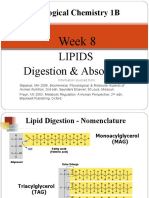
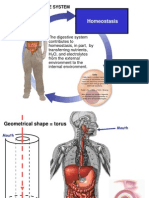
Potrebbero piacerti anche
- Fast Facts pour les patients: Les troubles d'oxydation des acides gras à chaîne longueDa EverandFast Facts pour les patients: Les troubles d'oxydation des acides gras à chaîne longueNessuna valutazione finora
- Fast Facts: Long-Chain Fatty Acid Oxidation Disorders for PatientsDa EverandFast Facts: Long-Chain Fatty Acid Oxidation Disorders for PatientsNessuna valutazione finora
- Lipid Metabolism L3+4Documento50 pagineLipid Metabolism L3+4bgj9cddvxhNessuna valutazione finora
- BCH 206 CholesterolDocumento3 pagineBCH 206 CholesterolNewtonNessuna valutazione finora
- Cholesterol FinalDocumento55 pagineCholesterol FinalSobha MatthewNessuna valutazione finora
- Digestion of Lipids. Transport Forms of LipidsDocumento91 pagineDigestion of Lipids. Transport Forms of LipidsAhmed FtsNessuna valutazione finora
- Lipo Protein MetDocumento32 pagineLipo Protein MetMilkoo sabaNessuna valutazione finora
- Physiotherapy (Post.) Lipid Metabolism DR - Amal BadrDocumento35 paginePhysiotherapy (Post.) Lipid Metabolism DR - Amal BadrthestaffforpediatricptNessuna valutazione finora
- Lipid Metabolism - (V) - Cholesterol and LipoproteinsDocumento35 pagineLipid Metabolism - (V) - Cholesterol and Lipoproteinslightning proNessuna valutazione finora
- Biochem Q.B III Sess Mar 2020Documento39 pagineBiochem Q.B III Sess Mar 2020Jinal Jyoti JoshiNessuna valutazione finora
- Lipid MetabolismDocumento17 pagineLipid MetabolismMiran El-MaghrabiNessuna valutazione finora
- Abetalipoproteinemia Powerpoint ReportDocumento43 pagineAbetalipoproteinemia Powerpoint Reportpurpleflurp23Nessuna valutazione finora
- Carnitine DeficiencyDocumento10 pagineCarnitine DeficiencyPam ElaNessuna valutazione finora
- Case 1Documento10 pagineCase 1DevianaNessuna valutazione finora
- Sirera, DyslipidemiaDocumento68 pagineSirera, Dyslipidemiarsirera6409Nessuna valutazione finora
- Cholesterol SphingolipidsDocumento31 pagineCholesterol SphingolipidsMaryam KNessuna valutazione finora
- Lipid MetabolismDocumento15 pagineLipid MetabolismMUTHONI IRERINessuna valutazione finora
- Checklist: A. B. C. D. eDocumento18 pagineChecklist: A. B. C. D. eHoanNessuna valutazione finora
- Metabolism of CholesterolDocumento17 pagineMetabolism of CholesterolAvhishek PatelNessuna valutazione finora
- Cholesterol Synthesis, Transport, and ExcretionDocumento36 pagineCholesterol Synthesis, Transport, and ExcretionabcdshNessuna valutazione finora
- Study 2 - ComprehensiveDocumento47 pagineStudy 2 - ComprehensivepeytonNessuna valutazione finora
- Metabolism of Lipids And: LipoproteinsDocumento50 pagineMetabolism of Lipids And: LipoproteinsRichoco SapoetraNessuna valutazione finora
- Energy Metabolism1Documento8 pagineEnergy Metabolism1edimulyanaNessuna valutazione finora
- SGL 7,8Documento30 pagineSGL 7,8hasan.hmu2004Nessuna valutazione finora
- Lipid Metabolism Dental and PhysiotherapyDocumento43 pagineLipid Metabolism Dental and PhysiotherapyNada Atef KoraitemNessuna valutazione finora
- Energy Production 2. Energy Storage 3. Energy UtilizationDocumento14 pagineEnergy Production 2. Energy Storage 3. Energy UtilizationLALITH SAI KNessuna valutazione finora
- Metabolism of LipidsDocumento37 pagineMetabolism of LipidsSafura IjazNessuna valutazione finora
- CB - Unit4 - Regulation of Lipid Metabolism and Ketone BodiesDocumento27 pagineCB - Unit4 - Regulation of Lipid Metabolism and Ketone BodiesBoomi BoomiNessuna valutazione finora
- Lec 6 Fed State PDFDocumento52 pagineLec 6 Fed State PDFrajeshNessuna valutazione finora
- Metabolism of LipidsDocumento98 pagineMetabolism of LipidsLyra GetNessuna valutazione finora
- Atherosclerosis: Reporter: Collera, Charissa Constantino, Venice Clemena, Adnan Coronel, Romeo Cordova, KarlaDocumento73 pagineAtherosclerosis: Reporter: Collera, Charissa Constantino, Venice Clemena, Adnan Coronel, Romeo Cordova, Karlaprecious_bustosNessuna valutazione finora
- Farmakoterapi DislipidemiaDocumento52 pagineFarmakoterapi DislipidemialisaNessuna valutazione finora
- Tenp DPT Biochemistry: DR Pius KiptemburDocumento13 pagineTenp DPT Biochemistry: DR Pius KiptemburGerald Limo Arap ChebiiNessuna valutazione finora
- Chapter 26Documento33 pagineChapter 26raul sinatoNessuna valutazione finora
- Overview of Metab and Carbo 1Documento16 pagineOverview of Metab and Carbo 1Jubilee Christiene AngNessuna valutazione finora
- L06 LipoproteinsDocumento23 pagineL06 LipoproteinsAbdullahi NorNessuna valutazione finora
- Lipid MetabolismDocumento21 pagineLipid MetabolismTooba GhummanNessuna valutazione finora
- Lipoproteins: SummaryDocumento3 pagineLipoproteins: SummaryEzekiel Morena RiveraNessuna valutazione finora
- HyperlipidemiaDocumento9 pagineHyperlipidemiaua6546830Nessuna valutazione finora
- Cholesterol Metabolism 2021Documento43 pagineCholesterol Metabolism 2021Eniola abdullahi AduagbaNessuna valutazione finora
- B314 AssignmentDocumento23 pagineB314 AssignmentPalesa NtsekalleNessuna valutazione finora
- Lipids Digestion and AbsorptionDocumento34 pagineLipids Digestion and Absorptionglenn johnstonNessuna valutazione finora
- LipidsDocumento13 pagineLipidsalianaNessuna valutazione finora
- Pharmacology :MIT:Lees:8Documento8 paginePharmacology :MIT:Lees:8Louis FortunatoNessuna valutazione finora
- Chem 113 Lipid ActivityDocumento3 pagineChem 113 Lipid ActivityAlliah MendozaNessuna valutazione finora
- Adrenal GlandDocumento46 pagineAdrenal GlandhSANNessuna valutazione finora
- Metabolism Summary Ch25Documento8 pagineMetabolism Summary Ch25Victor EkutaNessuna valutazione finora
- LIPID METABOLISM Notes PDFDocumento8 pagineLIPID METABOLISM Notes PDFToga BrandonNessuna valutazione finora
- Chapter 16 - Lipid MetabolismDocumento8 pagineChapter 16 - Lipid MetabolismadambridgerNessuna valutazione finora
- Cholesterol Synthesis, Transport, and Excretion: Abdul Salam M Sofro Faculty of Medicine YARSI UniversityDocumento39 pagineCholesterol Synthesis, Transport, and Excretion: Abdul Salam M Sofro Faculty of Medicine YARSI UniversitySofie Hanafiah NuruddhuhaNessuna valutazione finora
- An Overview of Lipid MetabolismDocumento19 pagineAn Overview of Lipid Metabolismshanto.tn98Nessuna valutazione finora
- 1 HendersonDocumento74 pagine1 Hendersonbballchamp10Nessuna valutazione finora
- Chronic Liver DiseaseDocumento12 pagineChronic Liver DiseaseamitNessuna valutazione finora
- Introduction To Enzymes Practical Handout For 2nd Year MBBSDocumento7 pagineIntroduction To Enzymes Practical Handout For 2nd Year MBBSIMDCBiochemNessuna valutazione finora
- Carbohydrate MetabolismDocumento4 pagineCarbohydrate MetabolismMuhammad Naveed SheikhNessuna valutazione finora
- 3rd Module 1Documento44 pagine3rd Module 1KARTHIK VINODNessuna valutazione finora
- Metabolic Functions of The LiverDocumento3 pagineMetabolic Functions of The LiverAizat KamalNessuna valutazione finora
- Ketone Bodies MetabolismDocumento3 pagineKetone Bodies MetabolismCristhian LozanoNessuna valutazione finora
- Biochemical Functions of LiverDocumento94 pagineBiochemical Functions of LiverMi PatelNessuna valutazione finora
- LIPDocumento28 pagineLIPsamar yousif mohamedNessuna valutazione finora
- Lecture 1 761 Pediatric Dentistry2011Documento42 pagineLecture 1 761 Pediatric Dentistry2011KgerbNessuna valutazione finora
- Document3 Spaced PDFDocumento17 pagineDocument3 Spaced PDFKgerbNessuna valutazione finora
- Lecture 3 761 Pediatric Dentistry 2011Documento45 pagineLecture 3 761 Pediatric Dentistry 2011KgerbNessuna valutazione finora
- Lecture 4 761 Pediataric Dentistry 2011Documento41 pagineLecture 4 761 Pediataric Dentistry 2011KgerbNessuna valutazione finora
- Ethics Notes PDFDocumento2 pagineEthics Notes PDFKgerbNessuna valutazione finora
- Lecture 2 761 Pediatric Dentistry 2011Documento33 pagineLecture 2 761 Pediatric Dentistry 2011KgerbNessuna valutazione finora
- Acute Pericarditis and Cardiac TamponadeDocumento49 pagineAcute Pericarditis and Cardiac TamponadesajidmuNessuna valutazione finora
- Document3 Spaced PDFDocumento17 pagineDocument3 Spaced PDFKgerbNessuna valutazione finora
- Old Midterm Q'SDocumento1 paginaOld Midterm Q'SKgerbNessuna valutazione finora
- GI Hormones PDFDocumento20 pagineGI Hormones PDFKgerb100% (1)
- Initial AssessmentDocumento6 pagineInitial AssessmentKgerbNessuna valutazione finora
- Cell Injury, Adaptation and DeathDocumento4 pagineCell Injury, Adaptation and DeathKgerbNessuna valutazione finora
- Implantprotocol-2013 WvuDocumento10 pagineImplantprotocol-2013 WvuKgerbNessuna valutazione finora
- DHR PDFDocumento3 pagineDHR PDFKgerbNessuna valutazione finora
- Dental HistoryDocumento2 pagineDental HistoryKgerbNessuna valutazione finora
- Document3 Spaced PDFDocumento17 pagineDocument3 Spaced PDFKgerbNessuna valutazione finora
- Digestion 1Documento59 pagineDigestion 1KgerbNessuna valutazione finora
- Department Performance Assessments Eligibility Criteria Jr. Benchmarks Performance/Assessments (Sr's Only) Sr. BenchmarksDocumento4 pagineDepartment Performance Assessments Eligibility Criteria Jr. Benchmarks Performance/Assessments (Sr's Only) Sr. BenchmarksKgerbNessuna valutazione finora
- Dental HistoryDocumento2 pagineDental HistoryKgerbNessuna valutazione finora
- CH 19 PDFDocumento2 pagineCH 19 PDFKgerbNessuna valutazione finora
- CH 20 PDFDocumento4 pagineCH 20 PDFKgerbNessuna valutazione finora
- CH 23 - Nervous System PDFDocumento2 pagineCH 23 - Nervous System PDFKgerbNessuna valutazione finora
- CH 16 PDFDocumento2 pagineCH 16 PDFKgerbNessuna valutazione finora
- CH 17 PDFDocumento4 pagineCH 17 PDFKgerbNessuna valutazione finora
- CH 14 PDFDocumento2 pagineCH 14 PDFKgerbNessuna valutazione finora
- GI Diseases (Excluding Oral Cavity) Esophagus StomachDocumento2 pagineGI Diseases (Excluding Oral Cavity) Esophagus StomachKgerbNessuna valutazione finora
- CH 14 PDFDocumento2 pagineCH 14 PDFKgerbNessuna valutazione finora
- CH 13 PDFDocumento3 pagineCH 13 PDFKgerbNessuna valutazione finora
- CH 11 PDFDocumento3 pagineCH 11 PDFKgerbNessuna valutazione finora
- Test Bank For Personal Nutrition 9th EditionDocumento32 pagineTest Bank For Personal Nutrition 9th Editionthomasburgessptrsoigxdk100% (21)
- Camellia Sinensis: Common NamesDocumento27 pagineCamellia Sinensis: Common NamesMohamad Saadatian SipanNessuna valutazione finora
- Understanding The Link Between Diabetes and Heart HealthDocumento3 pagineUnderstanding The Link Between Diabetes and Heart HealthDr Shashi BhushanNessuna valutazione finora
- Effect of Ethanol Leaf Extract of Chromolaena Odorata On Lipid Profile of Streptozotocin Induced Diabetic Wistar Albino RatsDocumento9 pagineEffect of Ethanol Leaf Extract of Chromolaena Odorata On Lipid Profile of Streptozotocin Induced Diabetic Wistar Albino RatsPUBLISHER JOURNALNessuna valutazione finora
- Appropriate UsSGLT2s and GLP-1RAs With Insulin To Reduce CVD Risk in Patients With Diabetes ArticleDocumento13 pagineAppropriate UsSGLT2s and GLP-1RAs With Insulin To Reduce CVD Risk in Patients With Diabetes ArticlenesredeNessuna valutazione finora
- Lesson 3 Notes Keto MasterclassDocumento7 pagineLesson 3 Notes Keto Masterclass2readNessuna valutazione finora
- Carbon Disulfide: I. Physical and Chemical PropertiesDocumento10 pagineCarbon Disulfide: I. Physical and Chemical PropertiesAditya ShrivastavaNessuna valutazione finora
- Activity 17 Clinical ChemistryDocumento53 pagineActivity 17 Clinical ChemistryKendrix Aguiluz FlorendoNessuna valutazione finora
- Cholecystolithiasis Case StudyDocumento70 pagineCholecystolithiasis Case StudyIrize DenagaNessuna valutazione finora
- Bhadoriya 2018Documento27 pagineBhadoriya 2018Gopal ReddyNessuna valutazione finora
- MNT 2 Case StudyDocumento14 pagineMNT 2 Case Studyapi-339312954Nessuna valutazione finora
- Cardiovascular DiseaseDocumento22 pagineCardiovascular DiseaseSunday JamesNessuna valutazione finora
- Apobt: Nacl Diluent 9%Documento3 pagineApobt: Nacl Diluent 9%Yunita RapaNessuna valutazione finora
- Automated LDL Cholesterol - Dimension - Rev D DXDCM 09008b838085cac0-1508285940764Documento15 pagineAutomated LDL Cholesterol - Dimension - Rev D DXDCM 09008b838085cac0-1508285940764Rizka Diana PutriNessuna valutazione finora
- Carbs & LipidsDocumento366 pagineCarbs & LipidsSafa ToqirNessuna valutazione finora
- COPD Significantly Increases Cerebral and Cardiovascular Events in HypertensivesDocumento8 pagineCOPD Significantly Increases Cerebral and Cardiovascular Events in HypertensivesAndreea MoalesNessuna valutazione finora
- Penatalaksanaan Dislipidemia Statin Sebagai Salah Satu Modalitas TerapiDocumento21 paginePenatalaksanaan Dislipidemia Statin Sebagai Salah Satu Modalitas TerapiDissa Naratania HantraNessuna valutazione finora
- Pratap Sanchetee: 23. Scientific View On FastingDocumento20 paginePratap Sanchetee: 23. Scientific View On Fastinglaiba ansariNessuna valutazione finora
- Variability in The Control of Type 2 Diabetes in Primary Care and Its Association With Hospital Admissions For Vascular Events. The APNA Study.Documento12 pagineVariability in The Control of Type 2 Diabetes in Primary Care and Its Association With Hospital Admissions For Vascular Events. The APNA Study.franciscoNessuna valutazione finora
- CP-67, Viraj Khand, Gomti Nagar, Lucknow - 226 010Documento9 pagineCP-67, Viraj Khand, Gomti Nagar, Lucknow - 226 010himanshu rockNessuna valutazione finora
- Troxerutin Is A Semisynthesis Compounds Based On RutinDocumento18 pagineTroxerutin Is A Semisynthesis Compounds Based On RutinSebastian NemethNessuna valutazione finora
- Transport in Blood Are Abnormal. Lipids AreDocumento4 pagineTransport in Blood Are Abnormal. Lipids AreAndi Marwan ZulkhaidirNessuna valutazione finora
- LFS 233:module 5 Fats: College of Health Sciences Integrated Sciences DepartmentDocumento65 pagineLFS 233:module 5 Fats: College of Health Sciences Integrated Sciences DepartmentHussain Ali100% (1)
- Aterogenik IndeksDocumento4 pagineAterogenik IndeksHizbulloh Putra AdiNessuna valutazione finora
- Zocor Application ApprovalDocumento216 pagineZocor Application ApprovalMohamed OmerNessuna valutazione finora
- Coronary Artery Diseases - Ilham PDocumento43 pagineCoronary Artery Diseases - Ilham PKeputrian FKUPNessuna valutazione finora
- 99fbcd52 Da4b 4be2 Bfd8 789c8ffdc7eeDocumento7 pagine99fbcd52 Da4b 4be2 Bfd8 789c8ffdc7eeG D HEALTH CARENessuna valutazione finora
- Lipids and LipoproteinsDocumento5 pagineLipids and LipoproteinsMica BernardoNessuna valutazione finora
- Antilipidemic DrugsDocumento34 pagineAntilipidemic Drugschelle_morales260% (1)
- Does Family History of Obesity, Cardiovascular, and Metabolic Diseases Influence Onset and Severity of Childhood ObesityDocumento6 pagineDoes Family History of Obesity, Cardiovascular, and Metabolic Diseases Influence Onset and Severity of Childhood ObesityTony Miguel Saba SabaNessuna valutazione finora