Potrebbero piacerti anche
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Chinese Paper Cutting Work SheetDocumento4 pagineChinese Paper Cutting Work Sheet黃梓Nessuna valutazione finora
- FinalDocumento18 pagineFinalAkash LadNessuna valutazione finora
- Epreuve Anglais EG@2022Documento12 pagineEpreuve Anglais EG@2022Tresor SokoudjouNessuna valutazione finora
- DFo 2 1Documento15 pagineDFo 2 1Donna HernandezNessuna valutazione finora
- PE MELCs Grade 3Documento4 paginePE MELCs Grade 3MARISSA BERNALDONessuna valutazione finora
- Mosharaf HossainDocumento2 pagineMosharaf HossainRuhul RajNessuna valutazione finora
- BARUDocumento53 pagineBARUhueuaNessuna valutazione finora
- Bajaj Vs Hero HondaDocumento63 pagineBajaj Vs Hero HondaHansini Premi100% (1)
- Virtual Assets Act, 2022Documento18 pagineVirtual Assets Act, 2022Rapulu UdohNessuna valutazione finora
- EN 50122-1 January 2011 Corrientes RetornoDocumento81 pagineEN 50122-1 January 2011 Corrientes RetornoConrad Ziebold VanakenNessuna valutazione finora
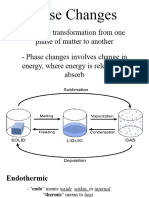
- General Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveDocumento19 pagineGeneral Chemistry 2 Q1 Lesson 5 Endothermic and Exotheric Reaction and Heating and Cooling CurveJolo Allexice R. PinedaNessuna valutazione finora
- Civ Beyond Earth HotkeysDocumento1 paginaCiv Beyond Earth HotkeysExirtisNessuna valutazione finora
- Siemens Rapidlab 248, 348, 840, 845, 850, 855, 860, 865: Reagents & ControlsDocumento2 pagineSiemens Rapidlab 248, 348, 840, 845, 850, 855, 860, 865: Reagents & ControlsJuan Carlos CrespoNessuna valutazione finora
- Gomez-Acevedo 2010 Neotropical Mutualism Between Acacia and Pseudomyrmex Phylogeny and Divergence TimesDocumento16 pagineGomez-Acevedo 2010 Neotropical Mutualism Between Acacia and Pseudomyrmex Phylogeny and Divergence TimesTheChaoticFlameNessuna valutazione finora
- EVOM ManualDocumento2 pagineEVOM ManualHouston WhiteNessuna valutazione finora
- Understanding PTS Security PDFDocumento37 pagineUnderstanding PTS Security PDFNeon LogicNessuna valutazione finora
- 2 To 20 Years - Girls Stature-For-Age and Weight-For-Age PercentilesDocumento1 pagina2 To 20 Years - Girls Stature-For-Age and Weight-For-Age PercentilesRajalakshmi Vengadasamy0% (1)
- HepaDocumento1 paginaHepasenthilarasu5100% (1)
- Chapter 2Documento8 pagineChapter 2Fahmia MidtimbangNessuna valutazione finora
- DION IMPACT 9102 SeriesDocumento5 pagineDION IMPACT 9102 SeriesLENEEVERSONNessuna valutazione finora
- Data Asimilasi Untuk PemulaDocumento24 pagineData Asimilasi Untuk PemulaSii Olog-olog PlonkNessuna valutazione finora
- Bank Danamon Organization ChartDocumento4 pagineBank Danamon Organization ChartFaisal Agus NugrahaNessuna valutazione finora
- List of Some Common Surgical TermsDocumento5 pagineList of Some Common Surgical TermsShakil MahmodNessuna valutazione finora
- Dakua Makadre PresentationDocumento12 pagineDakua Makadre PresentationEli Briggs100% (1)
- 3 Diversion&CareDocumento2 pagine3 Diversion&CareRyan EncomiendaNessuna valutazione finora
- Codan Rubber Modern Cars Need Modern Hoses WebDocumento2 pagineCodan Rubber Modern Cars Need Modern Hoses WebYadiNessuna valutazione finora
- EAC Inquiry SDCDocumento9 pagineEAC Inquiry SDCThe Sustainable Development Commission (UK, 2000-2011)Nessuna valutazione finora
- Wallem Philippines Shipping Inc. v. S.R. Farms (Laxamana)Documento2 pagineWallem Philippines Shipping Inc. v. S.R. Farms (Laxamana)WENDELL LAXAMANANessuna valutazione finora
- Lab Report SBK Sem 3 (Priscilla Tuyang)Documento6 pagineLab Report SBK Sem 3 (Priscilla Tuyang)Priscilla Tuyang100% (1)
- Facebook: Daisy BuchananDocumento5 pagineFacebook: Daisy BuchananbelenrichardiNessuna valutazione finora