Potrebbero piacerti anche
- ENCCEJA 2019 Ensino Médio PDFDocumento76 pagineENCCEJA 2019 Ensino Médio PDFPrincipe LogunedeNessuna valutazione finora
- Afetivograma em BrancoDocumento1 paginaAfetivograma em BrancotaniataubateNessuna valutazione finora
- Manual de Normas, Rotinas e POP - S de Enfermaem Do CAPSi RECRIAR (09.12) .19Documento185 pagineManual de Normas, Rotinas e POP - S de Enfermaem Do CAPSi RECRIAR (09.12) .19Maria Fernanda Alencar100% (1)
- Conceitos Mobilização e Manipulação Articular:: Professor Dr. Elias PortoDocumento36 pagineConceitos Mobilização e Manipulação Articular:: Professor Dr. Elias PortoRicardo PontesNessuna valutazione finora
- Uma Cor Chamada FLICTSDocumento8 pagineUma Cor Chamada FLICTSJuliana Maria CarvalhoNessuna valutazione finora
- Base Exercícios ResistidosDocumento31 pagineBase Exercícios ResistidosFrancisco JuliãoNessuna valutazione finora
- CHECCHIA. SOUZA JR. LIMA. (Orgs.) Por Uma Psicanálise Revolucionária. Otto GrossDocumento271 pagineCHECCHIA. SOUZA JR. LIMA. (Orgs.) Por Uma Psicanálise Revolucionária. Otto GrossFelipe Araujo67% (3)
- Síndrome Do ImpactoDocumento3 pagineSíndrome Do ImpactoSheila SilvaNessuna valutazione finora
- Avaliação Psicológica Pré-Cirurgia Bariátrica - A Experiência Dos PacientesDocumento9 pagineAvaliação Psicológica Pré-Cirurgia Bariátrica - A Experiência Dos PacientesMaria Gorete Arruda Oliveira PSICOLOGIANessuna valutazione finora
- Declaração Comparecimento Bloco A4Documento4 pagineDeclaração Comparecimento Bloco A4DORINELMA AlbuquerqueNessuna valutazione finora
- Atividade 23 - Descritor 13 - Grupo 3Documento8 pagineAtividade 23 - Descritor 13 - Grupo 3Juliana Maria CarvalhoNessuna valutazione finora
- Atividade 26 - Trabalhando Com Música - A Bailarina - Grupo 3 PDFDocumento2 pagineAtividade 26 - Trabalhando Com Música - A Bailarina - Grupo 3 PDFJuliana Maria CarvalhoNessuna valutazione finora
- Atividade 25 - Descritores 2 7 e 8 - Grupo 3 PDFDocumento10 pagineAtividade 25 - Descritores 2 7 e 8 - Grupo 3 PDFJuliana Maria CarvalhoNessuna valutazione finora
- Atividade 18 Descritor 12 Parte 1 Grupo 3Documento21 pagineAtividade 18 Descritor 12 Parte 1 Grupo 3Juliana Maria CarvalhoNessuna valutazione finora
- Comprovante Do Pagamento 1/2Documento2 pagineComprovante Do Pagamento 1/2Juliana Maria CarvalhoNessuna valutazione finora
- Direitos AutoraisDocumento1 paginaDireitos AutoraisJuliana Maria CarvalhoNessuna valutazione finora
- Atividade 26 - Trabalhando Com Música - A Bailarina - Grupo 3Documento2 pagineAtividade 26 - Trabalhando Com Música - A Bailarina - Grupo 3Juliana Maria CarvalhoNessuna valutazione finora
- Orientações para Disciplinas EADDocumento34 pagineOrientações para Disciplinas EADIlma AlmeidaNessuna valutazione finora
- Cognição e Novas TecnologiasDocumento18 pagineCognição e Novas TecnologiasJuliana Maria CarvalhoNessuna valutazione finora
- A Relevancia Do Designer Instrucional PTDocumento16 pagineA Relevancia Do Designer Instrucional PTJuliana Maria CarvalhoNessuna valutazione finora
- Pretto Educação e ComunicaçãoDocumento12 paginePretto Educação e ComunicaçãoJuliana Maria CarvalhoNessuna valutazione finora
- Abordagens EnsinoDocumento8 pagineAbordagens EnsinoJuliana Maria CarvalhoNessuna valutazione finora
- A Massa Crítica e A InternetDocumento4 pagineA Massa Crítica e A InternetJuliana Maria CarvalhoNessuna valutazione finora
- Qual A Importância de Trabalhar Com Os Alunos A Educação para o ConsumoDocumento2 pagineQual A Importância de Trabalhar Com Os Alunos A Educação para o ConsumoJuliana Maria CarvalhoNessuna valutazione finora
- Sistema Econômico Mundial - Gênese e Alcance de Um Conceito - Theotônio Dos SantosDocumento26 pagineSistema Econômico Mundial - Gênese e Alcance de Um Conceito - Theotônio Dos SantosJuliana Maria CarvalhoNessuna valutazione finora
- A Educação para A Economia CriativaDocumento4 pagineA Educação para A Economia CriativaJuliana Maria CarvalhoNessuna valutazione finora
- A Educação para A Economia CriativaDocumento4 pagineA Educação para A Economia CriativaJuliana Maria CarvalhoNessuna valutazione finora
- Conhecimento Aprendizagem e TecnologiasDocumento4 pagineConhecimento Aprendizagem e TecnologiasJuliana Maria CarvalhoNessuna valutazione finora
- Livro Biossegurança Editora SagahDocumento298 pagineLivro Biossegurança Editora SagahAvanúziaNessuna valutazione finora
- Política de Saúde Sobre HIVAIDS-1 PDFDocumento12 paginePolítica de Saúde Sobre HIVAIDS-1 PDFSuilan Moreira FerreiraNessuna valutazione finora
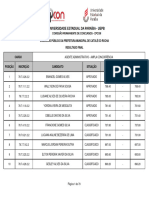
- Resultado Final Catole Do RochaDocumento79 pagineResultado Final Catole Do RochadgvcxfNessuna valutazione finora
- Cvi Argentina 3145965Documento4 pagineCvi Argentina 3145965ivo Machado SilvaNessuna valutazione finora
- VejaDocumento12 pagineVejaKELLYNessuna valutazione finora
- Vídeo H. Pneumotórax Hipertensivo e Vídeo Da Punção Descompressiva de Urgência - Dr. Herlon - Apresentação PDFDocumento15 pagineVídeo H. Pneumotórax Hipertensivo e Vídeo Da Punção Descompressiva de Urgência - Dr. Herlon - Apresentação PDFMariana BergerNessuna valutazione finora
- Termos e Condicoes 1 Dia - 230605 - 212959Documento9 pagineTermos e Condicoes 1 Dia - 230605 - 212959Equipa tribŌleoNessuna valutazione finora
- Ficha de Preparação Final - Trabalho III - Solução de Cloridrato de PropranololDocumento3 pagineFicha de Preparação Final - Trabalho III - Solução de Cloridrato de PropranololLuísa SilvaNessuna valutazione finora
- Apostila Visão e AudiçãoDocumento2 pagineApostila Visão e AudiçãocamilaNessuna valutazione finora
- Mapa Integrado - Segur - 91-2024Documento3 pagineMapa Integrado - Segur - 91-2024admcavaliniassessoriaNessuna valutazione finora
- Desgravada Finalissimia 20 - 21Documento411 pagineDesgravada Finalissimia 20 - 21Sérgio Daniel SilvaNessuna valutazione finora
- Proposta Adesão Seguro ITAUDocumento4 pagineProposta Adesão Seguro ITAUAlexandre SalesNessuna valutazione finora
- AULA PR TICA 01 - Imunologia CL Nica 27-09-2023Documento4 pagineAULA PR TICA 01 - Imunologia CL Nica 27-09-2023Thamires GabrielaNessuna valutazione finora
- Atividade Presencial para ConsultaDocumento9 pagineAtividade Presencial para ConsultaNatreFe50% (2)
- Análise Do Comportamento Aplicada Ao Autismo e Atuação Socialmente Responsável No BrasilDocumento16 pagineAnálise Do Comportamento Aplicada Ao Autismo e Atuação Socialmente Responsável No Brasilraphael guilherme costaNessuna valutazione finora
- Aula 1 - Introdução Ao Gerenciamento de EnfermagemDocumento32 pagineAula 1 - Introdução Ao Gerenciamento de EnfermagemPriscila M. V. de OliveiraNessuna valutazione finora
- Revista Edicao n21 PDFDocumento16 pagineRevista Edicao n21 PDFCarmely AlvesNessuna valutazione finora
- RDC 23 Anvisa Resolução 2012Documento7 pagineRDC 23 Anvisa Resolução 2012WalterNessuna valutazione finora
- RDC 11 2012Documento12 pagineRDC 11 2012spirotessNessuna valutazione finora
- CaninosDocumento5 pagineCaninosLuis Filipi RodriguesNessuna valutazione finora
- 25 Dicas Sobre A Qualidade Do ArDocumento34 pagine25 Dicas Sobre A Qualidade Do Arpedro_arsenioNessuna valutazione finora