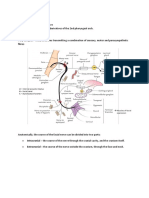
Potrebbero piacerti anche
- Pathophysiology of StrokeDocumento11 paginePathophysiology of StrokeAgronaSlaughterNessuna valutazione finora
- Pathophysiology of Stroke 2Documento12 paginePathophysiology of Stroke 2AgronaSlaughterNessuna valutazione finora
- Khoshnam2017 Article PathogenicMechanismsFollowingIDocumento20 pagineKhoshnam2017 Article PathogenicMechanismsFollowingILovin BiancaNessuna valutazione finora
- Patofisiologi StrkeDocumento14 paginePatofisiologi StrkeTeuku ILza Nanta SatiaNessuna valutazione finora
- Pa Tho Physiology of Traumatic Brain Injury2Documento22 paginePa Tho Physiology of Traumatic Brain Injury2n2s_nqNessuna valutazione finora
- Jurnal PDFDocumento9 pagineJurnal PDFFerdy Arif FadhilahNessuna valutazione finora
- Fisiopatología de La Isquemia Cerebral FocalDocumento10 pagineFisiopatología de La Isquemia Cerebral FocalshilacorinaNessuna valutazione finora
- Brain Injury After Cardiac Arrest PDocumento22 pagineBrain Injury After Cardiac Arrest PStrangerIn ParadiseNessuna valutazione finora
- Molecular Pathophysiology of StrokeDocumento10 pagineMolecular Pathophysiology of StrokeKahubbi Fatimah Wa'aliyNessuna valutazione finora
- Practice of NCC Hypoxic-Ischemic Encephalopathy in AdultsDocumento24 paginePractice of NCC Hypoxic-Ischemic Encephalopathy in AdultsAna Rica Santiago Navarra-CruzNessuna valutazione finora
- WIKI Cardio Vascular AccidentDocumento2 pagineWIKI Cardio Vascular AccidentAldrin CerdaNessuna valutazione finora
- Brain Injury Following Cardiac Arrest: Pathophysiology For Neurocritical CareDocumento10 pagineBrain Injury Following Cardiac Arrest: Pathophysiology For Neurocritical CareDianis DiNessuna valutazione finora
- Neurometabolic CascadeDocumento8 pagineNeurometabolic CascadeTim BonineNessuna valutazione finora
- Hyperglicemia in Stroke TreatmentDocumento13 pagineHyperglicemia in Stroke TreatmentChiquitaPutriVaniaRauNessuna valutazione finora
- Sub 1 Chir VascDocumento9 pagineSub 1 Chir Vascste fanNessuna valutazione finora
- Perspective of Synaptic Protection After Post-Infarction Treatment With StatinsDocumento9 paginePerspective of Synaptic Protection After Post-Infarction Treatment With StatinsDrGasnasNessuna valutazione finora
- Pathophysiology, Bioelectricidad, Review Stroke TINS99Documento7 paginePathophysiology, Bioelectricidad, Review Stroke TINS99Carla Saramy MachillandaNessuna valutazione finora
- Se FisiopatoDocumento28 pagineSe Fisiopatocska398Nessuna valutazione finora
- Ischemic StrokeDocumento8 pagineIschemic StrokeRinavi Adrin RirinNessuna valutazione finora
- Stroke: BackgroundDocumento10 pagineStroke: Backgroundjhay-r reyesNessuna valutazione finora
- Administering Corticosteroids in Neurologic Diseases3724Documento12 pagineAdministering Corticosteroids in Neurologic Diseases3724Carmen BritoNessuna valutazione finora
- Essential Update: New Tool For Predicting Renal Failure or Death in RhabdomyolysisDocumento12 pagineEssential Update: New Tool For Predicting Renal Failure or Death in RhabdomyolysisAndaru Kusuma PrajaNessuna valutazione finora
- Acute Stroke ArticleDocumento10 pagineAcute Stroke ArticlenesredeNessuna valutazione finora
- Clinic Trăilă of StemDocumento22 pagineClinic Trăilă of StemRadu AvramNessuna valutazione finora
- Epilepsy AssignmentDocumento58 pagineEpilepsy Assignmentnasir khanNessuna valutazione finora
- Cerebral Edema and Its Management - PMCDocumento1 paginaCerebral Edema and Its Management - PMCMohamed RawyNessuna valutazione finora
- FisiopatologiaDeLaLesionCerebral PDFDocumento10 pagineFisiopatologiaDeLaLesionCerebral PDFmagdalenaNessuna valutazione finora
- Cell InjuryDocumento8 pagineCell Injuryshalu abNessuna valutazione finora
- RESUME STROKE and SPINAL CORD INJURYDocumento8 pagineRESUME STROKE and SPINAL CORD INJURYYopy OctavianaNessuna valutazione finora
- 97 312 PDFDocumento5 pagine97 312 PDFdupuytrenNessuna valutazione finora
- Dapus 2Documento8 pagineDapus 2Beirnes Fernando Sembiring MNessuna valutazione finora
- ZORA Rudiger 2Documento25 pagineZORA Rudiger 2Carlos TrellesNessuna valutazione finora
- Intensive Care Treatment in Traumatic Brain Injury: The Epidemiology of Head TraumaDocumento6 pagineIntensive Care Treatment in Traumatic Brain Injury: The Epidemiology of Head TraumaDwi Utari JNessuna valutazione finora
- Neurochemical Cascade of ConcussionDocumento16 pagineNeurochemical Cascade of ConcussionJoan Choi100% (1)
- Beta-Block The Septic Heart 2010Documento5 pagineBeta-Block The Septic Heart 2010Ivann FloresNessuna valutazione finora
- Qin, Et Al. 2022Documento29 pagineQin, Et Al. 2022Ignacio BarbieriNessuna valutazione finora
- 2019 - Brain Functional Reserve in The Context of Neuroplasticity After StrokeDocumento10 pagine2019 - Brain Functional Reserve in The Context of Neuroplasticity After StrokeEl Tal RuleiroNessuna valutazione finora
- Neuroinflammatory Mechanisms in Ischemic Stroke: Focus On Cardioembolic Stroke, Background, and Therapeutic ApproachesDocumento33 pagineNeuroinflammatory Mechanisms in Ischemic Stroke: Focus On Cardioembolic Stroke, Background, and Therapeutic ApproachesSonia P SNessuna valutazione finora
- 3 I Ischamia InjuryDocumento50 pagine3 I Ischamia InjuryKartikey MishraNessuna valutazione finora
- Cell Injury: Dr. Sanjiv Kumar Assistant Professor, Deptt. of Pathology, BVC, PatnaDocumento32 pagineCell Injury: Dr. Sanjiv Kumar Assistant Professor, Deptt. of Pathology, BVC, PatnaAjinkya JadhaoNessuna valutazione finora
- Cell Injury, Adaptation and Death: HST.035 Spring 2003Documento30 pagineCell Injury, Adaptation and Death: HST.035 Spring 2003leksis09Nessuna valutazione finora
- Cell Injury - Afeef NazzalDocumento11 pagineCell Injury - Afeef NazzalFiras AnayaNessuna valutazione finora
- Bioquimica de La Isquemia y Reperfusion ArticuloDocumento35 pagineBioquimica de La Isquemia y Reperfusion ArticulofelipeNessuna valutazione finora
- Primary and Secondary InjuriesDocumento8 paginePrimary and Secondary Injuriesstella pangestikaNessuna valutazione finora
- Serum Electrolyte Derangements in Patients With Traumatic Brain InjuryDocumento3 pagineSerum Electrolyte Derangements in Patients With Traumatic Brain InjuryMartha OktaviaNessuna valutazione finora
- Perfusion CerebralDocumento10 paginePerfusion CerebralDrpoNessuna valutazione finora
- Shock: Focal PointDocumento15 pagineShock: Focal PointAndrea Carolina Rojas MendezNessuna valutazione finora
- Cerebral EdemaDocumento5 pagineCerebral EdemaAdilah AdeebNessuna valutazione finora
- Neurorestoration StrokeDocumento11 pagineNeurorestoration StrokeHastrina MailaniNessuna valutazione finora
- Fisiopatología de La Isquemia Cerebral Focal: Related PapersDocumento11 pagineFisiopatología de La Isquemia Cerebral Focal: Related PapersjansNessuna valutazione finora
- Cerebral EdemaDocumento6 pagineCerebral EdemaantonopoulosalNessuna valutazione finora
- Edema Serebri MJAFI 2003Documento6 pagineEdema Serebri MJAFI 2003Suryadi P. Dwipayana, Ida BgsNessuna valutazione finora
- Cell InjuryDocumento41 pagineCell InjuryMuhammad HuzaifaNessuna valutazione finora
- A Review On Cerebral Ischemic Stroke Mandeep Pundir, Manjinder Kour, Darshdeep SinghDocumento11 pagineA Review On Cerebral Ischemic Stroke Mandeep Pundir, Manjinder Kour, Darshdeep SinghParamjeet Kaur Paramedical SciencesNessuna valutazione finora
- A StrokeDocumento6 pagineA StrokeCrizzy GuiaoNessuna valutazione finora
- Reperfusion Injury: From Wikipedia, The Free EncyclopediaDocumento20 pagineReperfusion Injury: From Wikipedia, The Free EncyclopediaGenji TakiyaNessuna valutazione finora
- Basic Neurochemistry: Principles of Molecular, Cellular, and Medical NeurobiologyDa EverandBasic Neurochemistry: Principles of Molecular, Cellular, and Medical NeurobiologyNessuna valutazione finora
- Ischemic and Traumatic Brain and Spinal Cord Injuries: Mechanisms and Potential TherapiesDa EverandIschemic and Traumatic Brain and Spinal Cord Injuries: Mechanisms and Potential TherapiesNessuna valutazione finora
- PD 626 CasesDocumento5 paginePD 626 CasesOlan Dave LachicaNessuna valutazione finora
- Risk Management: Manoj VermaDocumento61 pagineRisk Management: Manoj Vermamverma1284Nessuna valutazione finora
- Muscle Grading and Testing ProceduresDocumento5 pagineMuscle Grading and Testing Proceduresshodhganga100% (1)
- Musculoskeletal Assessment ChecklistDocumento2 pagineMusculoskeletal Assessment ChecklistThe Blue and Gold RvdNessuna valutazione finora
- Diabetes Examination: General Appearance/InspectionDocumento5 pagineDiabetes Examination: General Appearance/InspectionNadia SalwaniNessuna valutazione finora
- Atlas of Approaches in Neurosurgery - J.FischerDocumento7 pagineAtlas of Approaches in Neurosurgery - J.FischerZdravko HeinrichNessuna valutazione finora
- Válvula 2 Stage Step StopDocumento34 pagineVálvula 2 Stage Step Stopmichelin25Nessuna valutazione finora
- Jet Bandsaw Manual - 414457Documento28 pagineJet Bandsaw Manual - 414457jfdcjhfgfglNessuna valutazione finora
- PDLDocumento2 paginePDLKathleen Cay Medrano PanopioNessuna valutazione finora
- Cure & CareDocumento10 pagineCure & CareZee ShanNessuna valutazione finora
- The Facial Nerve NotesDocumento5 pagineThe Facial Nerve NotesSimphiwe CebisaNessuna valutazione finora
- Analysis of Pathological GaitDocumento27 pagineAnalysis of Pathological GaitRica Mae AlimarioNessuna valutazione finora
- Appendicular SkeletonDocumento65 pagineAppendicular Skeletonnanak00ciaraNessuna valutazione finora
- Electrical Safety GlovesDocumento4 pagineElectrical Safety GlovesAnton Indra SupriatnaNessuna valutazione finora
- Lilius v. Manila Railroad CompanyDocumento6 pagineLilius v. Manila Railroad CompanyERNEST ELACH ELEAZARNessuna valutazione finora
- Ani Full AnswerDocumento7 pagineAni Full AnswerIshu & Gattu's Toy ReviewNessuna valutazione finora
- Federal Tort Claims Act Claim For Damage, Injury or DeathDocumento2 pagineFederal Tort Claims Act Claim For Damage, Injury or DeathScott D. HughesNessuna valutazione finora
- Management of Acute Trauma: Samuel P. Carmichael II, Nathan T. Mowery, R. Shayn Martin, J. Wayne MeredithDocumento43 pagineManagement of Acute Trauma: Samuel P. Carmichael II, Nathan T. Mowery, R. Shayn Martin, J. Wayne Meredithsportillag1Nessuna valutazione finora
- Ligament Handbook 3rd EditionDocumento59 pagineLigament Handbook 3rd EditionWilliam KisseNessuna valutazione finora
- June 9, 2015 Central Wisconsin ShopperDocumento24 pagineJune 9, 2015 Central Wisconsin ShoppercwmediaNessuna valutazione finora
- Drill Press ManualDocumento25 pagineDrill Press ManualitalianjeremyNessuna valutazione finora
- H&S Quiz AnswersDocumento4 pagineH&S Quiz AnswersHashir Badar50% (2)
- Three Digit BCD Counter MC1453B Data SheetDocumento12 pagineThree Digit BCD Counter MC1453B Data Sheetmata1802Nessuna valutazione finora
- Wrist Hand Finger ExercisesDocumento5 pagineWrist Hand Finger ExercisesNicolae MihaliNessuna valutazione finora
- B20M3L5 - Neck InjuriesDocumento4 pagineB20M3L5 - Neck InjuriesBarda GulanNessuna valutazione finora
- CW El CuerpoDocumento2 pagineCW El CuerpovbllchckNessuna valutazione finora
- Physical AssessmentDocumento19 paginePhysical AssessmentMamerto Q. Flores IIINessuna valutazione finora
- Gamma3: Long Nail R2.0Documento52 pagineGamma3: Long Nail R2.0Aravind DesaiNessuna valutazione finora
- Deck and Pool Tips For A Safe Summer, Building Safety Month: Week 3Documento2 pagineDeck and Pool Tips For A Safe Summer, Building Safety Month: Week 3Michigan NewsNessuna valutazione finora
- 2015-16 Physical Education SyllabusDocumento6 pagine2015-16 Physical Education Syllabusapi-235053410Nessuna valutazione finora