Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- GESTALT REVPEW Volume 1 1997 Number 3Documento105 pagineGESTALT REVPEW Volume 1 1997 Number 3Anika BozovicNessuna valutazione finora
- Just Therapy: Warihi Campbell, Kiwi Tamasese & Charles Waldegrave (The Family Centre)Documento5 pagineJust Therapy: Warihi Campbell, Kiwi Tamasese & Charles Waldegrave (The Family Centre)sun angelaNessuna valutazione finora
- Blood Longitudinal (T1) and Transverse (T2) Relaxation TimeDocumento11 pagineBlood Longitudinal (T1) and Transverse (T2) Relaxation TimekingpintamNessuna valutazione finora
- Dissecting The Effects of Ischemia and Reperfusion On The Coronary Microcirculation in A Rat Model of Acute Myocardial InfarctionDocumento18 pagineDissecting The Effects of Ischemia and Reperfusion On The Coronary Microcirculation in A Rat Model of Acute Myocardial InfarctionkingpintamNessuna valutazione finora
- Cerebrovascular Reactivity Mapping Without Gas ChallengesDocumento7 pagineCerebrovascular Reactivity Mapping Without Gas ChallengeskingpintamNessuna valutazione finora
- Sequence: Respiratory-Triggered Inversion Recovery-HASTE (IRHASTE) (31) Sequence With 4Documento6 pagineSequence: Respiratory-Triggered Inversion Recovery-HASTE (IRHASTE) (31) Sequence With 4kingpintamNessuna valutazione finora
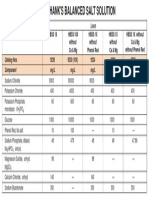
- HBSS FormulationDocumento1 paginaHBSS FormulationkingpintamNessuna valutazione finora
- Interferential Therapy Electrode Placement Techni-3 PDFDocumento9 pagineInterferential Therapy Electrode Placement Techni-3 PDFdivinaNessuna valutazione finora
- COPD LeafletDocumento2 pagineCOPD LeafletAN KnEeNessuna valutazione finora
- Aquilion One 640 LineasDocumento8 pagineAquilion One 640 LineasJussef M. ValverdeNessuna valutazione finora
- 00 Statistical Report 2016Documento248 pagine00 Statistical Report 2016wudthipanNessuna valutazione finora
- Trait Theory of PersonalityDocumento5 pagineTrait Theory of PersonalityBenitez GheroldNessuna valutazione finora
- Baclofen KemstroDocumento2 pagineBaclofen KemstroKristi WrayNessuna valutazione finora
- Ankle Proprioceptive ExercisesDocumento4 pagineAnkle Proprioceptive ExercisesDr. Krishna N. SharmaNessuna valutazione finora
- Ebpp Audit Tool - RevisedDocumento6 pagineEbpp Audit Tool - Revisedapi-259241412Nessuna valutazione finora
- Session 6: Alcohol and Drug Use: E. E. Mccollum, and K. H. RosenDocumento12 pagineSession 6: Alcohol and Drug Use: E. E. Mccollum, and K. H. RosenLidiaMaciasNessuna valutazione finora
- Annotated BibliographyDocumento4 pagineAnnotated Bibliographyapi-447388856Nessuna valutazione finora
- Copd and Cor PulmonalDocumento14 pagineCopd and Cor PulmonalAldi RafaelNessuna valutazione finora
- Nosocomial PneumoniaDocumento37 pagineNosocomial PneumoniaWinson ChitraNessuna valutazione finora
- Family Constellations An Innovative Systemic Pheno PDFDocumento9 pagineFamily Constellations An Innovative Systemic Pheno PDFFátimaA.SilvaNessuna valutazione finora
- 10 1002@14651858 CD004982 Pub6 PDFDocumento164 pagine10 1002@14651858 CD004982 Pub6 PDFghost_724Nessuna valutazione finora
- Một số nghiên cứu phác đồ diệt HpDocumento20 pagineMột số nghiên cứu phác đồ diệt HpPhạm Ngọc VănNessuna valutazione finora
- Pathology Of: NeoplasiaDocumento23 paginePathology Of: NeoplasiaBryan de GuzmanNessuna valutazione finora
- Dana Ullman HomeopathyDocumento4 pagineDana Ullman Homeopathydavid_moloneyNessuna valutazione finora
- Effects of Osteopathic Manipulative Treatment On Children With Plagiocephaly in The Context of Current Pediatric Practice - A Retrospective Chart Review StudyDocumento7 pagineEffects of Osteopathic Manipulative Treatment On Children With Plagiocephaly in The Context of Current Pediatric Practice - A Retrospective Chart Review StudymireliNessuna valutazione finora
- Creating A Productive Learning EnvironmentDocumento43 pagineCreating A Productive Learning EnvironmentDeSpirit Neverdies100% (1)
- Who Analgesic LadderDocumento55 pagineWho Analgesic LadderSyifa Nurul Asma'Nessuna valutazione finora
- Abnormal Illness BehaviorDocumento7 pagineAbnormal Illness Behavioransha2011p0% (1)
- Lester Et Al Cognitive ErrorsDocumento11 pagineLester Et Al Cognitive ErrorsДоникаNessuna valutazione finora
- Professional Nursing OrganizationDocumento4 pagineProfessional Nursing OrganizationCid Benedict PabalanNessuna valutazione finora
- Causes: Polycythemia VeraDocumento15 pagineCauses: Polycythemia VeraMaria VisitacionNessuna valutazione finora
- Anaphylaxis: Identification, Management and PreventionDocumento5 pagineAnaphylaxis: Identification, Management and PreventionÓscar A. Castejón CruzNessuna valutazione finora
- Asa Difficult Airway AlgorithmDocumento20 pagineAsa Difficult Airway AlgorithmspidymahiNessuna valutazione finora
- Periapical GranulomaDocumento6 paginePeriapical GranulomaEzza RiezaNessuna valutazione finora
- Susruthasamhitha Part 2Documento31 pagineSusruthasamhitha Part 2Dr Suvarna Nalapat100% (1)