Potrebbero piacerti anche
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Canadian Journal of DiabetesDocumento227 pagineCanadian Journal of DiabetesAnatolia MaresNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5795)
- Nab Technology PDFDocumento8 pagineNab Technology PDFalfox2000Nessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Irrational Use of Antibiotics and Role of The Pharmacist - An Insight From A Qualitative Study in New Delhi, India PDFDocumento6 pagineIrrational Use of Antibiotics and Role of The Pharmacist - An Insight From A Qualitative Study in New Delhi, India PDFalfox2000Nessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Pharmacist - S Role in Preventing Antibiotic Resistance PDFDocumento8 pagineThe Pharmacist - S Role in Preventing Antibiotic Resistance PDFalfox2000Nessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- INTERNATIONAL NONPROPRIETARY For Biological and Biotechnological Products PDFDocumento35 pagineINTERNATIONAL NONPROPRIETARY For Biological and Biotechnological Products PDFalfox2000Nessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Emetogenic Potential of Antineoplastic Agents PDFDocumento1 paginaEmetogenic Potential of Antineoplastic Agents PDFalfox2000Nessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Guidelines For Tablet Crushing and Administration Via Enteral Feeding Tubes PDFDocumento13 pagineGuidelines For Tablet Crushing and Administration Via Enteral Feeding Tubes PDFalfox2000Nessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Tema 3 - Inmune Responses To ImplantsDocumento18 pagineTema 3 - Inmune Responses To ImplantsIsrael GonzálezNessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- IN VITRO ANTIOXIDANT AND ANTIDIABETIC ACTIVITY of Plumeria PudicaDocumento10 pagineIN VITRO ANTIOXIDANT AND ANTIDIABETIC ACTIVITY of Plumeria PudicaTrinity PdplNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Topic 3 B Cell DevelopmentDocumento86 pagineTopic 3 B Cell DevelopmentPromita MajumdarNessuna valutazione finora
- Mark Scheme P1 and P2 KedahDocumento20 pagineMark Scheme P1 and P2 KedahRozaini OthmanNessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- Novel Concepts On Pregnancy Clocks and Alarms: Redundancy and Synergy in Human ParturitionDocumento26 pagineNovel Concepts On Pregnancy Clocks and Alarms: Redundancy and Synergy in Human ParturitionFedrik Monte Kristo LimbongNessuna valutazione finora
- Signal Transduction Pathways POGILDocumento6 pagineSignal Transduction Pathways POGILIsaiahGerardo100% (1)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Pediatric Pneumonia - Practice Essentials, Background, PathophysiologyDocumento1 paginaPediatric Pneumonia - Practice Essentials, Background, PathophysiologyShiferaw TesfayeNessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- C-Type Lectin Receptor-Induced NF-KB Activation in InnateDocumento8 pagineC-Type Lectin Receptor-Induced NF-KB Activation in InnateStefania CristinaNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Acupuncture Therapy in Ischemic Stroke RehabilitationDocumento12 pagineAcupuncture Therapy in Ischemic Stroke RehabilitationcocosinghNessuna valutazione finora
- (Mebooksfree Net) Adv&Pat&Cer&Str&Pat&Tri&Dwi&1stDocumento188 pagine(Mebooksfree Net) Adv&Pat&Cer&Str&Pat&Tri&Dwi&1stgabriellaNessuna valutazione finora
- Calcium and Klotho The Secret of ImmortalityDocumento1 paginaCalcium and Klotho The Secret of ImmortalityAgnes Vitta AngeliaNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- PS111 Brain & Behaviour Module 1: Psychobiology: Session 1 - IntroductionDocumento34 paginePS111 Brain & Behaviour Module 1: Psychobiology: Session 1 - IntroductionJayden TseNessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Molecular and Cellular Mechanisms of Biosignaling: Prof. Yong Tae KwonDocumento86 pagineMolecular and Cellular Mechanisms of Biosignaling: Prof. Yong Tae KwonJon QNessuna valutazione finora
- Abiotic Stress Response in Plants - Physiological Biochemical and Genetic PerspectivesDocumento358 pagineAbiotic Stress Response in Plants - Physiological Biochemical and Genetic PerspectivesCarolina Salazar ParraNessuna valutazione finora
- Major Catalogue Complet PDFDocumento237 pagineMajor Catalogue Complet PDFHervis FantiniNessuna valutazione finora
- Insulin Resistance PDFDocumento275 pagineInsulin Resistance PDFuzzal ahmedNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- MycomedicinalsDocumento51 pagineMycomedicinalslightningice100% (1)
- Chemical Signal: Hormone 1: Biosignaling (WEEK 2)Documento27 pagineChemical Signal: Hormone 1: Biosignaling (WEEK 2)Fenny Aulia SugianaNessuna valutazione finora
- Laila Jahn - Insulin - SignalingDocumento8 pagineLaila Jahn - Insulin - SignalingJennifer LarsonNessuna valutazione finora
- Clinical Nutrition - A Functional PerspectiveDocumento324 pagineClinical Nutrition - A Functional PerspectivelenaNessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Ulcerative Colitis Mucosal Transcriptomes Reveal Mitochondriopathy and Personalized Mechanisms Underlying Disease Severity and Treatment ResponseDocumento13 pagineUlcerative Colitis Mucosal Transcriptomes Reveal Mitochondriopathy and Personalized Mechanisms Underlying Disease Severity and Treatment ResponseAndriel CardosoNessuna valutazione finora
- CH 3 - Tissue RepairDocumento24 pagineCH 3 - Tissue RepairWesam AlTawilNessuna valutazione finora
- Recent Advances in NGF and Related Molecules: Laura Calzà Luigi Aloe Luciana Giardino EditorsDocumento302 pagineRecent Advances in NGF and Related Molecules: Laura Calzà Luigi Aloe Luciana Giardino EditorsPedro CastroNessuna valutazione finora
- Fibroblast Growth Factors: A Controlling Mechanism of Skin AgingDocumento8 pagineFibroblast Growth Factors: A Controlling Mechanism of Skin Agingnorma reyes contrerasNessuna valutazione finora
- CellDocumento14 pagineCellSyrian AsadNessuna valutazione finora
- Cell CommunicationDocumento32 pagineCell CommunicationMajd MerieNessuna valutazione finora
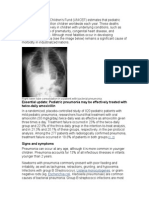
- Essential Update: Pediatric Pneumonia May Be Effectively Treated With Twice-Daily AmoxicillinDocumento54 pagineEssential Update: Pediatric Pneumonia May Be Effectively Treated With Twice-Daily AmoxicillinYostesara Maurena SantosaNessuna valutazione finora
- Do We Know JackDocumento20 pagineDo We Know JackAbdul-Ganiyu MannirNessuna valutazione finora
- ROS White Paper - 2015Documento21 pagineROS White Paper - 2015dzenitaNessuna valutazione finora
- Magnetic and Electromagnetic Field TherapyDocumento15 pagineMagnetic and Electromagnetic Field TherapyPaul VisageNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)