Potrebbero piacerti anche
- 301 LSP 11 Gen EngDocumento43 pagine301 LSP 11 Gen EngSuresh MgNessuna valutazione finora
- A11a BactVirusesDocumento17 pagineA11a BactVirusesSuresh MgNessuna valutazione finora
- Microbial and Non Microbial Contaminants of MilkDocumento5 pagineMicrobial and Non Microbial Contaminants of MilkSuresh MgNessuna valutazione finora
- Preparation For Diagnosing InfectionsDocumento26 paginePreparation For Diagnosing InfectionsSuresh MgNessuna valutazione finora
- PhagocytesDocumento16 paginePhagocytesSuresh MgNessuna valutazione finora
- Trna InternetDocumento37 pagineTrna InternetCandace HillNessuna valutazione finora
- Read The Newspaper Lately?: BIO 304 MicrobiologyDocumento18 pagineRead The Newspaper Lately?: BIO 304 MicrobiologySuresh MgNessuna valutazione finora
- 15 Eukaryotic Chromosomes Mitosis and MeiosisDocumento26 pagine15 Eukaryotic Chromosomes Mitosis and MeiosisSuresh MgNessuna valutazione finora
- Agricultural Biotechnology BiofertilizersDocumento38 pagineAgricultural Biotechnology BiofertilizersSuresh MgNessuna valutazione finora
- Biological Nitrogen Fixation and Biofertilizers Types and AdvantagesDocumento4 pagineBiological Nitrogen Fixation and Biofertilizers Types and AdvantagesSuresh MgNessuna valutazione finora
- Gopal - Preservation Techniques of MicroorganismDocumento14 pagineGopal - Preservation Techniques of MicroorganismSuresh MgNessuna valutazione finora
- Review Article On BiofertilizersDocumento17 pagineReview Article On BiofertilizersSuresh MgNessuna valutazione finora
- Cocclumn ChromatographyDocumento27 pagineCocclumn ChromatographySuresh MgNessuna valutazione finora
- Rdna TechnologyDocumento22 pagineRdna TechnologySuresh MgNessuna valutazione finora
- 391 110Documento7 pagine391 110Suresh MgNessuna valutazione finora
- Serial Dilution MethodDocumento3 pagineSerial Dilution Methoddraneesh75% (4)
- Enrichment Isolation Cultivation and Preservation of BacteriaDocumento56 pagineEnrichment Isolation Cultivation and Preservation of BacteriashuklagNessuna valutazione finora
- 119 928 1 PBDocumento4 pagine119 928 1 PBSuresh MgNessuna valutazione finora
- Electron MicroscopesDocumento5 pagineElectron MicroscopesSuresh MgNessuna valutazione finora
- Electron MicroscopesDocumento5 pagineElectron MicroscopesSuresh MgNessuna valutazione finora
- 2551 JMycologyMed17p.242-49Documento8 pagine2551 JMycologyMed17p.242-49Suresh MgNessuna valutazione finora
- Extraction and Purification of Antibacterial Metabolites From Actinomycetes Spp. Isolated From Soil SampleDocumento9 pagineExtraction and Purification of Antibacterial Metabolites From Actinomycetes Spp. Isolated From Soil SampleSuresh MgNessuna valutazione finora
- 513Documento4 pagine513Suresh MgNessuna valutazione finora
- Microscope Cleaning ProcedureDocumento1 paginaMicroscope Cleaning ProcedureSuresh MgNessuna valutazione finora
- Microscope 2 NotesDocumento3 pagineMicroscope 2 NotesSuresh MgNessuna valutazione finora
- Polymerase Chain ReactionDocumento13 paginePolymerase Chain ReactionSuresh MgNessuna valutazione finora
- Microscope 2 NotesDocumento3 pagineMicroscope 2 NotesSuresh MgNessuna valutazione finora
- Conventional Microscopy NotesDocumento1 paginaConventional Microscopy NotesSuresh MgNessuna valutazione finora
- 1 PCR History 2009aDocumento28 pagine1 PCR History 2009aSuresh MgNessuna valutazione finora
- Stock Culture and Its MaintenanceDocumento4 pagineStock Culture and Its MaintenanceSuresh Mg100% (2)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Anatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewDocumento7 pagineAnatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewPratistha ShauryaNessuna valutazione finora
- IV Fluid Management: Islam Awni Abu SamraDocumento41 pagineIV Fluid Management: Islam Awni Abu SamraIslam AwniNessuna valutazione finora
- Isometric Exercises SamsonDocumento30 pagineIsometric Exercises SamsonTom Hill78% (9)
- WV Nov 2023 First DraftDocumento8 pagineWV Nov 2023 First DraftMandlakhe HadebeNessuna valutazione finora
- Cassava Flour - Specification EAS 7402010Documento8 pagineCassava Flour - Specification EAS 7402010Suresh PatelNessuna valutazione finora
- Nursing Management of Pressure Ulcers in Adults PDFDocumento33 pagineNursing Management of Pressure Ulcers in Adults PDFAnindi Vivia MuktitamaNessuna valutazione finora
- Cysteamine - Full Profile - 170917Documento11 pagineCysteamine - Full Profile - 170917Walter MendozaNessuna valutazione finora
- Pediatric RehabDocumento32 paginePediatric RehabRainy DaysNessuna valutazione finora
- Biosep Short OverviewDocumento9 pagineBiosep Short OverviewMeyakorberNessuna valutazione finora
- Bio 14 NotesDocumento5 pagineBio 14 Noteshey lungNessuna valutazione finora
- 172-1610826745-Case ReportDocumento6 pagine172-1610826745-Case ReportBojan IlievNessuna valutazione finora
- Microsoft PowerPoint - Defense Mechanisms of The Gingiva (Compatibility MDocumento19 pagineMicrosoft PowerPoint - Defense Mechanisms of The Gingiva (Compatibility MDon Vigis AbrahamNessuna valutazione finora
- Contruccion de Vector de Expresion y Evaluación de Rendimiento de Biosimilar de Rituximab en CHODocumento7 pagineContruccion de Vector de Expresion y Evaluación de Rendimiento de Biosimilar de Rituximab en CHOSebastián Zapata LoaizaNessuna valutazione finora
- AP Bio: Chapter 12-The Cell Cycle Guided Reading Answer KeyDocumento4 pagineAP Bio: Chapter 12-The Cell Cycle Guided Reading Answer Keynikisepanj67% (6)
- TLC DLCDocumento67 pagineTLC DLCchandra shekharNessuna valutazione finora
- Kombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusDocumento9 pagineKombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusNeta AdzkiyaNessuna valutazione finora
- GPB 321 Crop Improvement Manual FinalDocumento71 pagineGPB 321 Crop Improvement Manual FinalSultan AhmadNessuna valutazione finora
- Chemistry FactsDocumento2 pagineChemistry FactsabcSDNessuna valutazione finora
- Dwnload Full Essentials of Understanding Psychology Canadian 5th Edition Feldman Test Bank PDFDocumento25 pagineDwnload Full Essentials of Understanding Psychology Canadian 5th Edition Feldman Test Bank PDFlogosdeviatorjx3b7i100% (9)
- Chapter 18Documento31 pagineChapter 18Ali KeresteciNessuna valutazione finora
- MD FTD SARS CoV2 Assay Spec Sheet 0820 FINAL - 1800000007432513Documento2 pagineMD FTD SARS CoV2 Assay Spec Sheet 0820 FINAL - 1800000007432513tranthien.gpvNessuna valutazione finora
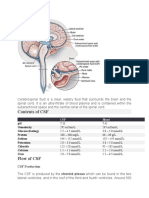
- Cerebrospinal FluidDocumento3 pagineCerebrospinal FluidlecturioNessuna valutazione finora
- Class 4 NSTSE PQP 10-Papers 2019-20 2-In-A4Documento80 pagineClass 4 NSTSE PQP 10-Papers 2019-20 2-In-A4Hatakishore BeheraNessuna valutazione finora
- Population Ecology: Aecc-I +3 1 YearDocumento32 paginePopulation Ecology: Aecc-I +3 1 YearAnita kumari SahuNessuna valutazione finora
- Article: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilDocumento15 pagineArticle: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilErikcsen Augusto RaimundiNessuna valutazione finora
- His T PathologyDocumento23 pagineHis T PathologygauravNessuna valutazione finora
- Pharmacogenetics: An Introduction: Alain Li-Wan-Po and Peter Farndon Introduce The Science ofDocumento3 paginePharmacogenetics: An Introduction: Alain Li-Wan-Po and Peter Farndon Introduce The Science ofDhuha NawabNessuna valutazione finora
- Bioreactor Types (Industrial Notes)Documento23 pagineBioreactor Types (Industrial Notes)anjaliNessuna valutazione finora
- Metabolism of XenobioticsDocumento84 pagineMetabolism of XenobioticsJNessuna valutazione finora
- Allegra 64R CentrifugeDocumento64 pagineAllegra 64R Centrifugeluroguita-1Nessuna valutazione finora