Potrebbero piacerti anche
- Nursing Practice Skills: Adult Intensive Care Unit PatientsDocumento10 pagineNursing Practice Skills: Adult Intensive Care Unit PatientsMona Doria67% (3)
- Aviation MagazinesDocumento46 pagineAviation Magazinesyogaga100% (1)
- Vein Type DepositDocumento7 pagineVein Type DepositHarisArmadiNessuna valutazione finora
- Desert Magazine 1957 DecemberDocumento44 pagineDesert Magazine 1957 Decemberdm1937Nessuna valutazione finora
- Global Projects Organisation: Material Specification For 316/316L and 6mo Austenitic Stainless SteelDocumento33 pagineGlobal Projects Organisation: Material Specification For 316/316L and 6mo Austenitic Stainless SteelThiyagarajan JayaramenNessuna valutazione finora
- Comparison of Several Methods For Effective Lipid Extraction From MicroalgaeDocumento3 pagineComparison of Several Methods For Effective Lipid Extraction From MicroalgaeRahul GogiaNessuna valutazione finora
- PH Meter Use and CalibrationDocumento10 paginePH Meter Use and CalibrationVlarick JongNessuna valutazione finora
- In Vitro Cell Culture Technique Protocol ManualDocumento48 pagineIn Vitro Cell Culture Technique Protocol ManualKavisa GhoshNessuna valutazione finora
- Biodiesel LabDocumento3 pagineBiodiesel LabGuilherme FriasNessuna valutazione finora
- Advance Probability and Statistics - 2014 - 2da Edición - Ottman PDFDocumento383 pagineAdvance Probability and Statistics - 2014 - 2da Edición - Ottman PDFAdan Graus RiosNessuna valutazione finora
- Imine ReductionDocumento4 pagineImine ReductionRatna Siti KhodijahNessuna valutazione finora
- GhostPeaks Part1Documento4 pagineGhostPeaks Part1Raga Bima100% (1)
- Sperm Cell: Specialised CellsDocumento2 pagineSperm Cell: Specialised CellsRhynnieNessuna valutazione finora
- Barge 180Ft Deck Load Capacity & Strength-Rev1Documento52 pagineBarge 180Ft Deck Load Capacity & Strength-Rev1Wahyu Codyr86% (7)
- Molecular Weight Cellulose EvansDocumento8 pagineMolecular Weight Cellulose Evansjulio070983Nessuna valutazione finora
- Production of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Filamentation-Suppressed Recombinant Escherichia ColiDocumento5 pagineProduction of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Filamentation-Suppressed Recombinant Escherichia Coliapi-3743140Nessuna valutazione finora
- Plant Macro-And Micronutrient MineralsDocumento5 paginePlant Macro-And Micronutrient Mineralsmanoj_rkl_07Nessuna valutazione finora
- DLVO TheoryDocumento22 pagineDLVO Theoryraj823Nessuna valutazione finora
- Ionic Liquids in Lipid Processing and Analysis: Opportunities and ChallengesDa EverandIonic Liquids in Lipid Processing and Analysis: Opportunities and ChallengesXuebing XuNessuna valutazione finora
- (Springer Laboratory) Prof. Sadao Mori, Dr. Howard G. Barth (Auth.) - Size Exclusion Chromatography-Springer-Verlag Berlin Heidelberg (1999)Documento235 pagine(Springer Laboratory) Prof. Sadao Mori, Dr. Howard G. Barth (Auth.) - Size Exclusion Chromatography-Springer-Verlag Berlin Heidelberg (1999)Luis Paulo BernardiNessuna valutazione finora
- Gel Filteration ChromatographyDocumento6 pagineGel Filteration ChromatographyAzka AsimNessuna valutazione finora
- Colloidal and Surface PhenomenaDocumento40 pagineColloidal and Surface PhenomenawynneralphNessuna valutazione finora
- Partition Coefficients and Their UsesDocumento92 paginePartition Coefficients and Their UsesquelenigNessuna valutazione finora
- Importance of Tris EDTADocumento15 pagineImportance of Tris EDTADarshana JuvekarNessuna valutazione finora
- ATCC - Animal Cell Culture Guide PDFDocumento39 pagineATCC - Animal Cell Culture Guide PDFAlier Guedes YeroNessuna valutazione finora
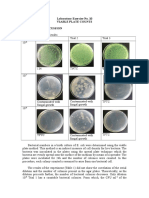
- Laboratory Exercise No. 10 Viable Plate Counts Results and DiscussionDocumento3 pagineLaboratory Exercise No. 10 Viable Plate Counts Results and Discussionvanessa olga100% (1)
- Experiment 1 (Introduction)Documento16 pagineExperiment 1 (Introduction)Msfaeza HanafiNessuna valutazione finora
- Exp 6 Citric Acid Production and EstimationDocumento9 pagineExp 6 Citric Acid Production and EstimationNikitaNessuna valutazione finora
- Ressitencia AntibioticosDocumento10 pagineRessitencia AntibioticosCarlota SoteloNessuna valutazione finora
- Answer HPLCDocumento3 pagineAnswer HPLCMuhammad Firdaus100% (1)
- Ftir and Gc-Fid CharacterizationDocumento8 pagineFtir and Gc-Fid CharacterizationIpeghan Otaraku100% (1)
- Membrane Research: Classic Origins and Current ConceptsDa EverandMembrane Research: Classic Origins and Current ConceptsA. L. Muggleton-HarrisNessuna valutazione finora
- Gel FiltrationDocumento123 pagineGel FiltrationZulfikri Asmardi RaufNessuna valutazione finora
- Regression Using SpssDocumento12 pagineRegression Using SpssArun RajanNessuna valutazione finora
- Yeast Protocols HandbookDocumento64 pagineYeast Protocols HandbookEmma StepinacNessuna valutazione finora
- Use of Micropippettor and SpectrophotometerDocumento6 pagineUse of Micropippettor and SpectrophotometerMichelleNessuna valutazione finora
- Size Exclusion ChromatographyDocumento15 pagineSize Exclusion ChromatographySumble AhmadNessuna valutazione finora
- Simulating Environment With Winogradsky Report 2Documento3 pagineSimulating Environment With Winogradsky Report 2Kai ChenNessuna valutazione finora
- Bet Surface Area Analysis of NanoparticlesDocumento11 pagineBet Surface Area Analysis of NanoparticlesKevin FungNessuna valutazione finora
- Downstream ProcessingDocumento4 pagineDownstream ProcessingVachaspatiMishraNessuna valutazione finora
- Prosonix - Sonocrystallization Proven Across Scale - 2009Documento10 pagineProsonix - Sonocrystallization Proven Across Scale - 2009Joshua JohnsonNessuna valutazione finora
- Nanowires - Recent AdvancesDocumento434 pagineNanowires - Recent AdvancesMohammad AliNessuna valutazione finora
- Biosepartaion Engineering: Ch.1: Bioseparation & Biological MaterialsDocumento89 pagineBiosepartaion Engineering: Ch.1: Bioseparation & Biological MaterialsAlex MaximusNessuna valutazione finora
- Ion ChromatographyDocumento2 pagineIon ChromatographyalexpharmNessuna valutazione finora
- Protein Extraction From Lupin Seeds-A Mathematical ModelDocumento11 pagineProtein Extraction From Lupin Seeds-A Mathematical Modeltbbdocument1s0% (1)
- Nucleosidic Phosphoramidite Synthesis Via PhosphitylationDocumento8 pagineNucleosidic Phosphoramidite Synthesis Via PhosphitylationClarence AG YueNessuna valutazione finora
- 2016 BCH223 PracticalsDocumento7 pagine2016 BCH223 PracticalsvictorNessuna valutazione finora
- The Influence of Conformational Isomerism On Drug ActionDocumento6 pagineThe Influence of Conformational Isomerism On Drug Actionhectorlope45Nessuna valutazione finora
- MTT Assay Sample ReportDocumento5 pagineMTT Assay Sample ReportAlexander VincentNessuna valutazione finora
- Isolation of DNA From Animal TissuesDocumento10 pagineIsolation of DNA From Animal TissuesAnura BandaraNessuna valutazione finora
- J. A. Radley-Examination and Analysis of Starch and Starch Products-Springer (1976)Documento222 pagineJ. A. Radley-Examination and Analysis of Starch and Starch Products-Springer (1976)KUKU100% (1)
- Biotechnology Lab Manual Offered To Iii Year B.Tech Chemical EngineeringDocumento34 pagineBiotechnology Lab Manual Offered To Iii Year B.Tech Chemical Engineeringanon_348923763Nessuna valutazione finora
- Intercalation Compounds of Graphite PDFDocumento186 pagineIntercalation Compounds of Graphite PDFchanchan88vn100% (1)
- Ion Exchange ChromatographyDocumento32 pagineIon Exchange Chromatographyவித்யா சேகர்Nessuna valutazione finora
- SpectrophotometryDocumento19 pagineSpectrophotometryabhinav_ramana100% (1)
- Exercise 1Documento13 pagineExercise 1Estephen FortelaNessuna valutazione finora
- Mini PrepDocumento6 pagineMini PrepWilson GomargaNessuna valutazione finora
- Aeration and AgitationDocumento9 pagineAeration and AgitationKasun Prasanna SilvaNessuna valutazione finora
- MTT AssayDocumento2 pagineMTT AssayHameedhaNessuna valutazione finora
- Separation by High-Speed Countercurrent ChromatographyDocumento17 pagineSeparation by High-Speed Countercurrent ChromatographyI'ah ZamzamiNessuna valutazione finora
- Fayaz Report On Challenges Associated With Sepn of Hcl-Water Azeotrope 04.08Documento27 pagineFayaz Report On Challenges Associated With Sepn of Hcl-Water Azeotrope 04.08fayaz5uin1234Nessuna valutazione finora
- Chemical Composition of Mung BeanDocumento8 pagineChemical Composition of Mung BeanEuler MiquilenaNessuna valutazione finora
- Experiment 37B-2 Spectroscopic Analysis of Dyes - More Than Pretty ColorsDocumento5 pagineExperiment 37B-2 Spectroscopic Analysis of Dyes - More Than Pretty ColorsHEEBA SATHYANessuna valutazione finora
- Clostridium Saccharobutylicum DSM 13864 Using Gelatinised Sago Starch As ADocumento9 pagineClostridium Saccharobutylicum DSM 13864 Using Gelatinised Sago Starch As AputrianneNessuna valutazione finora
- Experiment 5 ChromatographyDocumento3 pagineExperiment 5 ChromatographyJames Quan100% (2)
- Dextran and Related PolysaccharidesDocumento7 pagineDextran and Related PolysaccharidesEmadNessuna valutazione finora
- Fourier Transforms New Analytical Approaches and FTIR StrategiesDocumento509 pagineFourier Transforms New Analytical Approaches and FTIR StrategiesLuis David Cajavilca CutimancoNessuna valutazione finora
- Extraction of Native Protein From YeastDocumento5 pagineExtraction of Native Protein From YeastBrandon Lam100% (1)
- Evolution of Ecosystems - Terrestrial PDFDocumento4 pagineEvolution of Ecosystems - Terrestrial PDFmanoj_rkl_07Nessuna valutazione finora
- Ca Binding Proteins A0001347-001-000 PDFDocumento8 pagineCa Binding Proteins A0001347-001-000 PDFmanoj_rkl_07Nessuna valutazione finora
- 2,4-Thiazolidinedione As Antimicrobial and Cytotoxic AgentsDocumento10 pagine2,4-Thiazolidinedione As Antimicrobial and Cytotoxic Agentsmanoj_rkl_07Nessuna valutazione finora
- Evolution of Genome Organizn1699-001-000 PDFDocumento5 pagineEvolution of Genome Organizn1699-001-000 PDFmanoj_rkl_07Nessuna valutazione finora
- Ephrins: Ru Diger KleinDocumento6 pagineEphrins: Ru Diger Kleinmanoj_rkl_07Nessuna valutazione finora
- BMP Antags &nural Inducna0000805-001-000 PDFDocumento5 pagineBMP Antags &nural Inducna0000805-001-000 PDFmanoj_rkl_07Nessuna valutazione finora
- Evolution of Development A0001661-001-000 PDFDocumento4 pagineEvolution of Development A0001661-001-000 PDFmanoj_rkl_07Nessuna valutazione finora
- Calibration of A Transillumanator For UV Fixation of DNA To PDFDocumento2 pagineCalibration of A Transillumanator For UV Fixation of DNA To PDFmanoj_rkl_07Nessuna valutazione finora
- Heavy Metal Adaptation PDFDocumento4 pagineHeavy Metal Adaptation PDFmanoj_rkl_07Nessuna valutazione finora
- Electron Carriers PDFDocumento8 pagineElectron Carriers PDFmanoj_rkl_07100% (1)
- Cyanogenesis in Higher Plant and InsectsDocumento3 pagineCyanogenesis in Higher Plant and Insectsmanoj_rkl_07Nessuna valutazione finora
- Electroporation: Jac A NickoloffDocumento3 pagineElectroporation: Jac A Nickoloffmanoj_rkl_07Nessuna valutazione finora
- DNA Damage: Paul W DoetschDocumento7 pagineDNA Damage: Paul W Doetschmanoj_rkl_07Nessuna valutazione finora
- DNA Repair by Reversal of Damage PDFDocumento8 pagineDNA Repair by Reversal of Damage PDFmanoj_rkl_07100% (1)
- Flocyt Anal Nstem Cel in DevDocumento11 pagineFlocyt Anal Nstem Cel in Devmanoj_rkl_07Nessuna valutazione finora
- Forward-Cristopher Reeve PDFDocumento1 paginaForward-Cristopher Reeve PDFmanoj_rkl_07Nessuna valutazione finora
- Genetic Code Introduction PDFDocumento10 pagineGenetic Code Introduction PDFmanoj_rkl_07Nessuna valutazione finora
- FGF1Documento9 pagineFGF1manoj_rkl_07Nessuna valutazione finora
- Closteroviridae: Historical PerspectiveDocumento6 pagineClosteroviridae: Historical Perspectivemanoj_rkl_07Nessuna valutazione finora
- Immunologival Tolerance Therpeutic Induction PDFDocumento6 pagineImmunologival Tolerance Therpeutic Induction PDFmanoj_rkl_07Nessuna valutazione finora
- Phyllosphere PDFDocumento8 paginePhyllosphere PDFmanoj_rkl_07Nessuna valutazione finora
- Dideoxy Sequencing of DNA PDFDocumento16 pagineDideoxy Sequencing of DNA PDFmanoj_rkl_07Nessuna valutazione finora
- Dideoxy Sequencing of DNA PDFDocumento16 pagineDideoxy Sequencing of DNA PDFmanoj_rkl_07Nessuna valutazione finora
- Root Nodules (Rhizobium Legumes) PDFDocumento2 pagineRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07Nessuna valutazione finora
- Calibration of A Transillumanator For UV Fixation of DNA To PDFDocumento2 pagineCalibration of A Transillumanator For UV Fixation of DNA To PDFmanoj_rkl_07Nessuna valutazione finora
- Capillary Blotting of RNA and DNA Gels PDFDocumento3 pagineCapillary Blotting of RNA and DNA Gels PDFmanoj_rkl_07Nessuna valutazione finora
- Heavy Metal Adaptation PDFDocumento4 pagineHeavy Metal Adaptation PDFmanoj_rkl_07Nessuna valutazione finora
- Gertrude Belle Elion PDFDocumento1 paginaGertrude Belle Elion PDFmanoj_rkl_07Nessuna valutazione finora
- Terpenoids Lower PDFDocumento7 pagineTerpenoids Lower PDFmanoj_rkl_07Nessuna valutazione finora
- Architecture of HimalayasDocumento3 pagineArchitecture of HimalayasAndrea CaballeroNessuna valutazione finora
- Flood RateDocumento367 pagineFlood RatetpktpkNessuna valutazione finora
- Gas Permeability, Diffusivity and Solubility of Nitrogen, Helium, Methane, CO2, CH2O in Dense Polymeric Membranes Using A New On-Line Permeation ApparatusDocumento12 pagineGas Permeability, Diffusivity and Solubility of Nitrogen, Helium, Methane, CO2, CH2O in Dense Polymeric Membranes Using A New On-Line Permeation ApparatusCinthiaCevallosNessuna valutazione finora
- CHEM333 Syllabus 2020 2021Documento4 pagineCHEM333 Syllabus 2020 2021lina kwikNessuna valutazione finora
- DSE MC G11 G12 Equations Straight Lines 2023Documento6 pagineDSE MC G11 G12 Equations Straight Lines 2023ernestchan501Nessuna valutazione finora
- RH Fs Risk FactorsDocumento2 pagineRH Fs Risk FactorsfentroispNessuna valutazione finora
- TMJDocumento38 pagineTMJAnonymous 6kAmeZzINessuna valutazione finora
- Video Wall, Digital Signage & C'Type Serial NumberDocumento8 pagineVideo Wall, Digital Signage & C'Type Serial NumberAbhijit BarmanNessuna valutazione finora
- KIN-CN-STU-NW-0001 Puerto Real Towing Study of Kincardine 04Documento44 pagineKIN-CN-STU-NW-0001 Puerto Real Towing Study of Kincardine 04RUBEN BARTOLOME GARCIA100% (1)
- Esthetics and Shade Communication: A Practical Approach: Clinical ApplicationDocumento21 pagineEsthetics and Shade Communication: A Practical Approach: Clinical Applicationcatalin_adinaNessuna valutazione finora
- Chapter-01 Introduction: Sonadanga Residential Area (1st Phase)Documento17 pagineChapter-01 Introduction: Sonadanga Residential Area (1st Phase)MAFRID HAYDARNessuna valutazione finora
- Tugas Topic 4 Devi PermatasariDocumento8 pagineTugas Topic 4 Devi PermatasariMartinaNessuna valutazione finora
- Details of Placed Students in Academic Session 2022-23Documento10 pagineDetails of Placed Students in Academic Session 2022-23Rahul MishraNessuna valutazione finora
- Unit-3: Grid FrameworkDocumento44 pagineUnit-3: Grid FrameworkMUKESH KUMAR P 2019-2023 CSENessuna valutazione finora
- Ap Art and Design Drawing Sustained Investigation Samples 2019 2020 PDFDocumento102 pagineAp Art and Design Drawing Sustained Investigation Samples 2019 2020 PDFDominic SandersNessuna valutazione finora
- Hira - For Shot Blasting & Upto 2nd Coat of PaintingDocumento15 pagineHira - For Shot Blasting & Upto 2nd Coat of PaintingDhaneswar SwainNessuna valutazione finora
- Coulomb's Law and Electric Field Intensity: Engineering ElectromagneticsDocumento24 pagineCoulomb's Law and Electric Field Intensity: Engineering ElectromagneticsKenn SenadosNessuna valutazione finora
- Davao Region Slogan Reflective EssayDocumento4 pagineDavao Region Slogan Reflective EssayDonna Elaine OrdoñezNessuna valutazione finora
- Total Aircraft Washing System: Mudmaster Mudmaster MudliteDocumento2 pagineTotal Aircraft Washing System: Mudmaster Mudmaster MudliteAli Abu ShhiwaNessuna valutazione finora
- Post-Installed Rebars Design and Analysis: Singapore, 11.03.2016Documento44 paginePost-Installed Rebars Design and Analysis: Singapore, 11.03.2016SewGuanNessuna valutazione finora
- Philippine Airlines Reservation New Timings Dep - 230314 - 193643Documento7 paginePhilippine Airlines Reservation New Timings Dep - 230314 - 193643sophia buiserNessuna valutazione finora
- Report On RoboticsDocumento40 pagineReport On Roboticsangelcrystl4774Nessuna valutazione finora