Potrebbero piacerti anche
- Adult Infectious Disease Bulletpoints HandbookDa EverandAdult Infectious Disease Bulletpoints HandbookValutazione: 4.5 su 5 stelle4.5/5 (9)
- Ijmr 5 2 179 183Documento5 pagineIjmr 5 2 179 183Abhay KumarNessuna valutazione finora
- Molecular and Cellular Biology of Pathogenic TrypanosomatidsDa EverandMolecular and Cellular Biology of Pathogenic TrypanosomatidsNessuna valutazione finora
- Cefoxitin For AmpcDocumento6 pagineCefoxitin For AmpcmemossaNessuna valutazione finora
- Detection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.Documento6 pagineDetection of Genes TEM, OXA, SHV and CTX-M in 73 Clinical Isolates of Escherichia Coli Producers of Extended Spectrum Betalactamases and Determination of Their Susceptibility To Antibiotics.International Medical PublisherNessuna valutazione finora
- Alikhani 2014Documento6 pagineAlikhani 2014WahyunitadotokaNessuna valutazione finora
- Prevalence of Extended-Spectrum Beta-Lactamases Among Isolated Isolated From Blood Culture in A Tertiary Care HospitalDocumento4 paginePrevalence of Extended-Spectrum Beta-Lactamases Among Isolated Isolated From Blood Culture in A Tertiary Care HospitalMohammad K AlshomraniNessuna valutazione finora
- Antimicrobial Resistance and Molecular Characterization of Gene Cassettes From Class 1 Integrons in Pseudomonas Aeruginosa S PDFDocumento7 pagineAntimicrobial Resistance and Molecular Characterization of Gene Cassettes From Class 1 Integrons in Pseudomonas Aeruginosa S PDFValentina RondonNessuna valutazione finora
- Comparative Study of Cefoperazone Ceftazidime CeftDocumento3 pagineComparative Study of Cefoperazone Ceftazidime CeftAdityaNessuna valutazione finora
- ESBL Phenotipic PDFDocumento14 pagineESBL Phenotipic PDFGerardo J. Huerta RoblesNessuna valutazione finora
- Jurnal NCBIDocumento6 pagineJurnal NCBIPutri Nilam SariNessuna valutazione finora
- 4 C 566 A 70 Da 279 BCFDocumento8 pagine4 C 566 A 70 Da 279 BCFMazin AlmaziniNessuna valutazione finora
- RAPD AcinetoDocumento8 pagineRAPD AcinetokiflyNessuna valutazione finora
- Current Senerio of Antimicrobial Resistance Patterns of Pseudomonas Aeruginosa Clinical Isolates in East Uttar Pradesh, IndiaDocumento4 pagineCurrent Senerio of Antimicrobial Resistance Patterns of Pseudomonas Aeruginosa Clinical Isolates in East Uttar Pradesh, IndiaPraveen GautamNessuna valutazione finora
- Aztreonam ReviewDocumento10 pagineAztreonam Reviewncpscientist_2Nessuna valutazione finora
- 5 Psae Egypt 2014Documento9 pagine5 Psae Egypt 2014Khoa Vi Sinh BVCRNessuna valutazione finora
- Review B-Lactamase Production in Enterobacteriaceae: Review and Bench GuideDocumento14 pagineReview B-Lactamase Production in Enterobacteriaceae: Review and Bench GuideyomnayasminNessuna valutazione finora
- 22 1-S2.0-S0924857908002392-MainDocumento3 pagine22 1-S2.0-S0924857908002392-MainLookpear ShiiNessuna valutazione finora
- Paterson Et Al., 2021Documento3 paginePaterson Et Al., 2021Caio Bonfim MottaNessuna valutazione finora
- Art 5 PDFDocumento10 pagineArt 5 PDFpaola lopezNessuna valutazione finora
- Babol Jbums v17n8p19 enDocumento8 pagineBabol Jbums v17n8p19 enTendi SulaksoNessuna valutazione finora
- 3127-Article Text-25112-1-10-20131114Documento8 pagine3127-Article Text-25112-1-10-20131114guizah nourelhoudaNessuna valutazione finora
- Em Proteus Mirabilis em – Analysis of ADocumento6 pagineEm Proteus Mirabilis em – Analysis of Aranjani093Nessuna valutazione finora
- Yamano 2006 Occurrence of PER-1Documento6 pagineYamano 2006 Occurrence of PER-1WahyunitadotokaNessuna valutazione finora
- Multidrug Resistant On PseudomonasDocumento5 pagineMultidrug Resistant On PseudomonasMinh Nguyen HoangNessuna valutazione finora
- Antibiotic Impregnated Tablets For Screening Esbl and Ampc Beta LactamasesDocumento4 pagineAntibiotic Impregnated Tablets For Screening Esbl and Ampc Beta LactamasesIOSR Journal of PharmacyNessuna valutazione finora
- Role of The Mycobacterium Tuberculosis P55 Efflux Pump in Intrinsic Drug Resistance, Oxidative Stress Responses, and GrowthDocumento8 pagineRole of The Mycobacterium Tuberculosis P55 Efflux Pump in Intrinsic Drug Resistance, Oxidative Stress Responses, and GrowthAnnette GonzalezNessuna valutazione finora
- 2Documento4 pagine2Mohammad K AlshomraniNessuna valutazione finora
- tmp7DDD TMPDocumento6 paginetmp7DDD TMPFrontiersNessuna valutazione finora
- Cephalosporin Resistance in EnterobacteriacaeDocumento4 pagineCephalosporin Resistance in EnterobacteriacaeGIST (Gujarat Institute of Science & Technology)Nessuna valutazione finora
- Escherichia Coli in A Rural Medical College of Himachal PradeshDocumento5 pagineEscherichia Coli in A Rural Medical College of Himachal PradeshIOSR Journal of PharmacyNessuna valutazione finora
- Yang 2021Documento6 pagineYang 2021perencanaandinkesnaganrayaNessuna valutazione finora
- ESBLDocumento87 pagineESBLMona Mostafa100% (1)
- Molecular Characterization of Clinical Multidrug-Resistant Klebsiella Pneumoniae IsolatesDocumento5 pagineMolecular Characterization of Clinical Multidrug-Resistant Klebsiella Pneumoniae IsolatesSamman zakaNessuna valutazione finora
- 10.33073 - PJM 2022 023Documento6 pagine10.33073 - PJM 2022 023Andy SetiawanNessuna valutazione finora
- Genetic and Antibiotic Susceptibility Profiles of Drug - Resistant Acinetobacter Baumannii From Various Parts of LebanonDocumento9 pagineGenetic and Antibiotic Susceptibility Profiles of Drug - Resistant Acinetobacter Baumannii From Various Parts of LebanonInternational Medical PublisherNessuna valutazione finora
- Antimicrob. Agents Chemother.-2000-Bingen-1453-7Documento5 pagineAntimicrob. Agents Chemother.-2000-Bingen-1453-7WilliNessuna valutazione finora
- Zahra 2014Documento6 pagineZahra 2014WahyunitadotokaNessuna valutazione finora
- Molecular Epidemiologic Analysis of Enterococcus FaecalisDocumento8 pagineMolecular Epidemiologic Analysis of Enterococcus FaecalisoinkNessuna valutazione finora
- Inhibitor-Based Methods For Detection of Plasmid-Mediated Ampc - Lactamases in Klebsiella SPP., Escherichia Coli, and Proteus MirabilisDocumento5 pagineInhibitor-Based Methods For Detection of Plasmid-Mediated Ampc - Lactamases in Klebsiella SPP., Escherichia Coli, and Proteus MirabilisYanuar PrasetyoNessuna valutazione finora
- Art 03Documento6 pagineArt 03Luis MartinNessuna valutazione finora
- Guay2002 - Cefdinir An Advanced Generation Broad Spectrum Oral CephalosporinDocumento17 pagineGuay2002 - Cefdinir An Advanced Generation Broad Spectrum Oral CephalosporinTulus Java Rimbunan S.Nessuna valutazione finora
- Resistencia Ceftazidima-AvibactamDocumento4 pagineResistencia Ceftazidima-Avibactamjuan godinezNessuna valutazione finora
- Courvalin 1996Documento9 pagineCourvalin 1996Miy AichNessuna valutazione finora
- Antimicrobial Resistance in Campylobacter SPP Isolated FromDocumento3 pagineAntimicrobial Resistance in Campylobacter SPP Isolated FromEliud RománNessuna valutazione finora
- Porins, Efflux Pumps and Multidrug Resistance in Acinetobacter BaumanniiDocumento6 paginePorins, Efflux Pumps and Multidrug Resistance in Acinetobacter BaumanniiMaria MolinaNessuna valutazione finora
- Begum 2013Documento4 pagineBegum 2013WahyunitadotokaNessuna valutazione finora
- Dex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisDocumento6 pagineDex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisNona NonicaaNessuna valutazione finora
- ESBLDocumento2 pagineESBLCla ValenciaNessuna valutazione finora
- A Ten Disk Procedure For The Detection of Antibiotic Resistance in EnterobacteriacaeDocumento7 pagineA Ten Disk Procedure For The Detection of Antibiotic Resistance in EnterobacteriacaeNguyen Huu HienNessuna valutazione finora
- Antimicrobial Agents and Chemotherapy-2015-Pöppel-2508.fullDocumento7 pagineAntimicrobial Agents and Chemotherapy-2015-Pöppel-2508.fullEEDIEB Prof. Millton Marques CurvoNessuna valutazione finora
- 1998 CTX JacDocumento3 pagine1998 CTX JacSergey SidorenkoNessuna valutazione finora
- Karakterisasi Molekular Fragmen Gen MexbDocumento10 pagineKarakterisasi Molekular Fragmen Gen MexbHermanto MangandeNessuna valutazione finora
- Pseudomonas Aeruginosa - A Phenomenon of Bacterial ResistanceDocumento16 paginePseudomonas Aeruginosa - A Phenomenon of Bacterial ResistanceErinaa SetyoriniNessuna valutazione finora
- Outbreak of Carbapenem-Resistant Acinetobacter Baumannii Producing The Carbapenemase OXA-58 in TurkeyDocumento8 pagineOutbreak of Carbapenem-Resistant Acinetobacter Baumannii Producing The Carbapenemase OXA-58 in TurkeyDrashua AshuaNessuna valutazione finora
- Appl. Environ. Microbiol. 2001 Steidle 5761 70Documento10 pagineAppl. Environ. Microbiol. 2001 Steidle 5761 70Iulia IonNessuna valutazione finora
- 2011 PK-PD AntibiotikDocumento88 pagine2011 PK-PD AntibiotikUniatimelindaNessuna valutazione finora
- Aac 00642-17Documento8 pagineAac 00642-17Robert StryjakNessuna valutazione finora
- Incidence of Metallo Beta-Lactamase Producing Pseudomonas Aeruginosa in Clinical SamplesDocumento4 pagineIncidence of Metallo Beta-Lactamase Producing Pseudomonas Aeruginosa in Clinical SamplesvanishivshettyNessuna valutazione finora
- Xin 2016, Pfge&MlstDocumento8 pagineXin 2016, Pfge&MlstSarah KKCNessuna valutazione finora
- Cefalosporin Mnemonic 2015Documento3 pagineCefalosporin Mnemonic 2015Mari FereNessuna valutazione finora
- Here Is The Astro Chart You RequestedDocumento4 pagineHere Is The Astro Chart You RequestedMari FereNessuna valutazione finora
- LGC StadializareDocumento1 paginaLGC StadializareMari FereNessuna valutazione finora
- Orvosi LapokDocumento1 paginaOrvosi LapokMari FereNessuna valutazione finora
- Valori HBA1c in Mmoli Si %Documento1 paginaValori HBA1c in Mmoli Si %Mari FereNessuna valutazione finora
- Pachet Promo MicsoratDocumento1 paginaPachet Promo MicsoratMari FereNessuna valutazione finora
- Irene SheriDocumento2 pagineIrene SheriMari FereNessuna valutazione finora
- IeltsDocumento39 pagineIeltsMari FereNessuna valutazione finora
- IBR (Biochimie) CopiiDocumento4 pagineIBR (Biochimie) CopiiMari FereNessuna valutazione finora
- Multifibren UDocumento6 pagineMultifibren UMari Fere100% (1)
- IeltsDocumento1 paginaIeltsMari FereNessuna valutazione finora
- Lymekor DR BozsikDocumento1 paginaLymekor DR BozsikMari FereNessuna valutazione finora
- Chapter 24 HWDocumento5 pagineChapter 24 HWFola SolarinNessuna valutazione finora
- Richa Shrivastava Assistant Professor Department of Pharmacy BITS PilaniDocumento18 pagineRicha Shrivastava Assistant Professor Department of Pharmacy BITS PilaniCHOUGULE KISHOR SURYAKANTNessuna valutazione finora
- SOP 3.04 DNA Extraction From BloodDocumento4 pagineSOP 3.04 DNA Extraction From BloodMeetali GuptaNessuna valutazione finora
- Chapter 16 The Citric Acid Cycle: Multiple Choice QuestionsDocumento14 pagineChapter 16 The Citric Acid Cycle: Multiple Choice QuestionshielNessuna valutazione finora
- Escherichia Coli DH10BDocumento2 pagineEscherichia Coli DH10BSecc. 2 Marco Antonio Aviles RomeroNessuna valutazione finora
- Reviews: Epidemiology, Definition and Treatment of Complicated Urinary Tract InfectionsDocumento15 pagineReviews: Epidemiology, Definition and Treatment of Complicated Urinary Tract InfectionsPutu AdiNessuna valutazione finora
- Gent. Patho. Ento. MCQDocumento18 pagineGent. Patho. Ento. MCQvkrawani121Nessuna valutazione finora
- Anat MuscleDocumento164 pagineAnat Musclejohn delaNessuna valutazione finora
- Weekly Science P4 February 2021 PDFDocumento4 pagineWeekly Science P4 February 2021 PDFMs SinarNessuna valutazione finora
- Diagnosis and Management of Addiction and Other Mental DisordersDocumento208 pagineDiagnosis and Management of Addiction and Other Mental DisordersAhana MukherjeeNessuna valutazione finora
- 3 Carbohydrates' StructureDocumento33 pagine3 Carbohydrates' StructureDilan TeodoroNessuna valutazione finora
- 5 Enzymes: 0610 BiologyDocumento9 pagine5 Enzymes: 0610 BiologyAbdul HadiNessuna valutazione finora
- Gentics by Garima MamDocumento83 pagineGentics by Garima Mamgargmahek904Nessuna valutazione finora
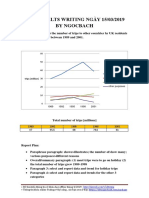
- Giai de Ielts Writing Ngay 150319 by NgocbachDocumento7 pagineGiai de Ielts Writing Ngay 150319 by NgocbachSedeaNessuna valutazione finora
- 6i. Male Reproduction - OnPRC Module 2-Student HandoutDocumento12 pagine6i. Male Reproduction - OnPRC Module 2-Student HandoutAkash ShawNessuna valutazione finora
- Cardiorespiratory EnduranceDocumento38 pagineCardiorespiratory EnduranceHilarion Vidal OlimpoNessuna valutazione finora
- Diagnosis Cure Treatment, Disease Prevention: Sources of Drug InformationDocumento11 pagineDiagnosis Cure Treatment, Disease Prevention: Sources of Drug InformationShyen100% (11)
- Full Download Test Bank For Evolutionary Psychology The New Science of The Mind 4 e 4th Edition David Buss PDF Full ChapterDocumento36 pagineFull Download Test Bank For Evolutionary Psychology The New Science of The Mind 4 e 4th Edition David Buss PDF Full Chapterrainbowenergy.t6op100% (19)
- Fagro 05 1183691Documento15 pagineFagro 05 1183691Maraza Choque Gisela JulyNessuna valutazione finora
- Stafford & Golightly - LSDDocumento102 pagineStafford & Golightly - LSDPicior de LemnNessuna valutazione finora
- TDZ For in Vitro Propagation of AlstroemeriaDocumento10 pagineTDZ For in Vitro Propagation of AlstroemeriaBrij Mohan SinghNessuna valutazione finora
- Dna DissertationDocumento6 pagineDna DissertationCustomCollegePaperUK100% (1)
- Syllabus For Soldier General Duty/Sol TDN (X) /sol TDN (Viii) PatternDocumento2 pagineSyllabus For Soldier General Duty/Sol TDN (X) /sol TDN (Viii) PatternAnooj KumarNessuna valutazione finora
- Cat's Lair - Christine FeehanDocumento462 pagineCat's Lair - Christine FeehanVivi Lara33% (3)
- AssignmentDocumento5 pagineAssignmentMELAKU ADIMASNessuna valutazione finora
- Yu 2022Documento11 pagineYu 2022FebbNessuna valutazione finora
- Fetal Skull PDFDocumento2 pagineFetal Skull PDFLaurieNessuna valutazione finora
- Dr. Ramakrishna Bag: Civil Engineering Dept NIT RourkelaDocumento17 pagineDr. Ramakrishna Bag: Civil Engineering Dept NIT RourkelaJon JimmyNessuna valutazione finora
- Compost Tea Brewing ManualDocumento91 pagineCompost Tea Brewing ManualJanetS14338100% (1)
- An Annotated Checklist of The Angiospermic Flora of Rajkandi Reserve Forest of Moulvibazar, BangladeshDocumento21 pagineAn Annotated Checklist of The Angiospermic Flora of Rajkandi Reserve Forest of Moulvibazar, BangladeshRashed KaramiNessuna valutazione finora
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDa EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNessuna valutazione finora
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDa EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDValutazione: 5 su 5 stelle5/5 (3)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDa EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionValutazione: 4 su 5 stelle4/5 (404)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDa EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityValutazione: 4 su 5 stelle4/5 (32)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDa EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeValutazione: 2 su 5 stelle2/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDa EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Da EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Valutazione: 3 su 5 stelle3/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDa EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedValutazione: 4.5 su 5 stelle4.5/5 (82)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDa EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsValutazione: 4 su 5 stelle4/5 (4)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisDa EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisValutazione: 4.5 su 5 stelle4.5/5 (42)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDa EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsValutazione: 5 su 5 stelle5/5 (1)
- The Obesity Code: Unlocking the Secrets of Weight LossDa EverandThe Obesity Code: Unlocking the Secrets of Weight LossValutazione: 4 su 5 stelle4/5 (6)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesDa EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesValutazione: 4.5 su 5 stelle4.5/5 (1412)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDa EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeValutazione: 4.5 su 5 stelle4.5/5 (254)
- The Marshmallow Test: Mastering Self-ControlDa EverandThe Marshmallow Test: Mastering Self-ControlValutazione: 4.5 su 5 stelle4.5/5 (60)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisDa EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisValutazione: 5 su 5 stelle5/5 (8)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Da EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Valutazione: 4.5 su 5 stelle4.5/5 (110)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4.5 su 5 stelle4.5/5 (6)
- To Explain the World: The Discovery of Modern ScienceDa EverandTo Explain the World: The Discovery of Modern ScienceValutazione: 3.5 su 5 stelle3.5/5 (51)
- Summary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedDa EverandSummary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedValutazione: 4 su 5 stelle4/5 (61)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDa EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryValutazione: 4 su 5 stelle4/5 (46)