Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
U2.Capitulo 7. Exitación Del Músculo Esquelético Transmisión Neuromuscular y Acoplamiento Exitación Contracción
Caricato da
Jaime Andres MontoyaTitolo originale
Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
U2.Capitulo 7. Exitación Del Músculo Esquelético Transmisión Neuromuscular y Acoplamiento Exitación Contracción
Caricato da
Jaime Andres MontoyaCopyright:
Formati disponibili
e
Excitacin del msculo esqueltico: transmisin neuromuscular y acoplamiento excitacin-contraccin
Transmisin de impulsos desde las terminaciones nerviosas a las fibras del msculo esqueltico: la unin neuromuscular
Las fibras del msculo esqueltico estn inervadas por fibras nerviosas mielinizadas grandes que se originan en las motoncuronas grandes de las astas anteriores de la mdula espinal. Como se ha sealado en el captulo 6. to das las fibras nerviosas, despus de entrar en el vientre muscular. normalmente se ramifican y estimulan entre tres y varios cientos ele fibras musculares esquelticas. Cada terminacin nerviosa forma una unin. denominada unin neuromuscular, con la fibra muscular cerca de su punto medio. El potencial de accin que se inicia en la fibra muscular por la seal nerviosa viaja en ambas direcciones hacia los extre mos de la fibra muscular. Con la excepcin de aproximadamente e12% de las fibras musculares. slo hay una unin de este tipo en cada fibra muscular.
de la unin neuromuscular: la placa motora terminal. La figu ra 7-1A y B muestra la unin ncuromuscular que forma una gran fibra nerviosa mie linizada con una fibra muscular esqueltica. La fibra nerviosa forma un complejo de Terminaciones nerviosas ramificadas que se invaginan en la superficie de la fibra muscular. pero que permanecen fuera de la membrana plasmtica de la misma. TOda la estructura se denomina placa motora terminal, Esta cubierta por una o ms clu las de Schwann que la aslan de los lquidos circundantes. La figura 7-1C muestra el esquema de una microfotografa electrnica de la unin entre una terminacin axnica nica y la membrana de una fibra muscular. La membrana invaginada se denomina gotiera sinptica o valle sinptico y el espacio que hay entre la terminacin y la membrana de la fibra se denomina espacio sinp tico o hendidura sinptica. Este espacio mide de 20 a 30 nanmetros de anchura. En el fondo de la gotiera hay numerosos pliegues ms pequeos de la membrana ele la fibra muscular denominados hendiduras subneurales. que aumentan mucho el rea su perficial en la que puede actuar el transmisor sinptico. En la terminacin axnica hay muchas mitocondrias que proporcionan trifosfato de adenosina (ATP), la fuente de energa que se utiliza para la sntesis del transmi sor excitador acetilcolina. La acetilcoJina. a su vez, excita a la membrana de la fibra muscular. La acetilcolina se sintetiza en el citoplasma de la terminacin. pero se ab sorbe rpidamente hacia el interior de muchas pequeas vesiculas sinpticas, de las que normalmente hay aproximadamente 300.000 en las terminaciones de una nica placa terminal. En el espacio sinptico hay grandes cantidades de la enzima acetil colinesterasa, que destruye la acctilcolina algunos milisegundos despus de que la hayan liberado las vesculas sinpticas,
Anatoma fisiolgica
Secrecin de acetilcolina por las terminaciones nerviosas
Cuando un impulso nervioso llega a la unin neuromuscular, se liberan aproxima damente 125 vesculas de acetilcolina desde las terminaciones hacia el espacio si nptico. Algunos de los detalles de este mecanismo se pueden ver en la figura 7-2.
85
86
Unidad11
Fisiologade la membrana,el nervio y el msculo
Clula de la telogla
Hendiduras subneurales
Diferentes imgenes de la placa motora terrninal. A. Corte longitu dinal a travs de la placa termi nal. B. Imagen de la superficie de la placa terminal. C. Aspecto en la microfotografla electrnica del punto de contacto entre una ni ca terminacin axnica y la mem brana de la fibra muscular. (Re producido a partir de Fawcett DW, segn la modificacin de Cou teaux R. in Bloom W, Fawcett DW: A Textbook of Histology, Phla delphia. WB Saunders, 1986.)
que muestra una imagen ampliada de un espacio sinpti co con la membrana neural por encima y la membrana muscular y sus hendiduras subneurales por debajo. En la superficie interna de la membrana neural hay barras densas lineales, que se muestran en seccin trans versal en la figura 7-2. A ambos lados de cada una de estas barras densas hay partculas proteicas que penetran en la membrana neural; son canales de calcio activados por el voltaje. Cuando un potencial de accin se propaga por la terminacin, estos canales se abren y permiten que iones calcio difundan desde el espacio sinptico hacia el interior de la terminacin nerviosa. Se piensa que a su vez los iones calcio ejercen una influencia de atraccin sobre las vesculas de acetilcolina, desplazndolas hacia la membra na neural adyacente a las barras densas. Las vesculas se fusionan con la membrana neural y vacan su acetilcolina ha cia el espacio sinptico mediante el proceso de exocitosis. Aunque algunos de los detalles que se han menciona do previamente son hipotticos, se sabe que el estmulo eficaz que produce la liberacin de acetilcolina desde las ve sculas es la entrada de iones calcio y que despus se vaca la acetilcolina desde las vesculas a travs de la membrana neural adyacente a las barras densas.
Puntos Membrana de liberacin neural
Vesculas
Hendidura subneural
Membrana muscular
figura
7_-_2J
Efecto de la acetilcolina sobre la membrana de la fibra muscular postsinptica para abrir canales inicos. La figu ra 7-2 tambin muestra muchos receptores de acetilcolina muy pequeos en la membrana de la fibra muscular; son ca nales inicos activados por acetilcolina, y estn localiza
dos casi totalmente cerca de las aberturas de las hendidu ras subneurales que estn inmediatamente debajo de las
Liberacin de acetilcolina desde las vesculas sinpticas en la membrana neural de la unin neuromuscular. Obsrvese la proxi midad de los puntos de liberacin de la membrana neuraJcon los receptores de acetilcolina de la membrana muscular. en las aber turas de las hendiduras subneurales.
zonas de las barras densas, donde se libera la acetilcolina hacia el espacio sinptico. Cada receptor es un complejo proteico que tiene un peso molecular total de 275.000. El complejo est forma-
Captulo 7
Excitacin del msculo esqueltico: transmisin neuromuscular y acoplamiento excitacin-contraccin
87
do por cinco subundades proteicas, dos protenas aLfa y una protena beta, una delta y una gamma. Estas molculas proteicas atraviesan la membrana. y estn dispuestas en crculo para formar un canal tubular, que se muestra en la figura 7-3.El canal permanece cerrado, como se ilustra en la seccinA de la figura, hasta que dos molculas de acetil colina se unen respectivamente a las dos subunidades pro teicas alfa. Esto produce un cambio conformacional que abre el canal, como se puede ver en la seccin B de la figura. El canal de acetilcolina abierto tiene un dimetro de aproximadamente 0,65 nm, que es lo suficientemente grande como para permitir que los iones positivos impor tantes (sodio [Na'], potasio [K+] y calcio lCa se mue van con facilidad a travs de la abertura. Por el contrario, los iones negativos, como los iones cloruro, no lo atravie san debido a las intensas cargas negativas de la abertura del canal que las repelen. En la prctica fluyen muchos ms iones sodio a travs de los canales de acetilcolina que de cualquier otro tipo, por dos motivos. Primero. slo hay dos iones positivos en
H])
concentraciones grandes: iones sodio en el Lquidoextra celular e iones potasio en ellfquido intracelular. Segundo, el potencial muy negativo del interior de la membrana muscular, de -80 a -90 IDV,arrastra los iones sodio de car ga positiva hacia el interior de la fibra. a la vez que impi de de manera simultnea la salida de los iones potasio de carga positiva cuando intentan pasar hacia el exterior. Como se muestra en la figura 7-38, el principal efecto de la apertura de loscanales activadospor la acetilcolinaes per mitir que grandes cantidades de iones sodio entren al in terior de la fibra, desplazando con ellos grandes nmeros de cargas positivas. Esto genera un cambio de potencial positivo local en la membrana de la fibra muscular. deno minado potencial de la pLaca terminal. A su vez. este po tencial de la placa terminal inicia un potencial de accin que se propaga a lo largo de la membrana muscular y de esta manera produce la contraccin muscular. Destruccin por la acetilcolinesterasa de la acetllcolina Ii berada. Una vez que se ha liberado hacia el espacio si nptico, la acetilcolina sigue activando los receptores de acetilcolina mientras persista en el espacio. Sin embargo. se elimina rpidamente por dos medios: 1) La mayor parte de la acetilcolina es destruida por la enzima acetitcolines terasa, que est unida principalmente a la capa esponjosa de tejido conjuntivo fino que llena el espacio sinptico entre la terminacin nerviosa prcsinptica y la membrana muscular postsinpca. 2) Una pequea cantidad de ace tilcolina difunde hacia el exterior del espacio sinptico y ya no est disponible para actuar sobre la membrana de la fibra muscular. El breve espacio de tiempo que permanece la acetilco lina en el espacio sinptico (algunos milisegundos como mucho) normalmente es suficiente para excitar la fibra muscular. Despus, la rpida eliminacin de la acetilcolina impide la reexcitacin muscular continuada despus de que la fibra muscular se haya recuperado de su potencial de accin inicial. Potencial de la placa terminal y excitacin de la fibra muscu lar esqueltica. La rpida entrada de iones sodio en la fi bra muscular cuando se abren los canales de acetilcolina hace que el potencial elctrico en el interior de la fibra en ]a zona local de la placa terminal aumente en direccin positiva hasta 50 a 75 mV, generando un potencial loca! denominado potencial de la placa terminal. Recurdese del captulo 5 que normalmente es suficiente un aumento sbito del potencial de la membrana nerviosa de ms de 20 a 30 mV para iniciar la apertura de cada vez ms cana les de sodio, iniciando de esta manera un potencial de ac cin en la membrana de la fibra muscular. La figura 7-4 muestra el principio del inicio del poten cial de accin por un potencial de La placa terminal. Esta fi gura muestra tres potenciales distintos de placa terminal. Los potenciales de la placa terminal A y C son demasiado dbiles para producir un potencial de accin, aunque sf producen cambios locales dbiles del voltaje de la placa terminal,como se muestra en la figura Porel contrario.el po tencial de la placa terminal B es mucho ms intenso y hace que se abra un nmero suficiente de canales de sodio, de modo que el efecto autorregcnerativo del flujo cada vez mayor de iones sodio hacia el interior de la fibra iniciaun po-
A
Na+
c::{ACh
8
Figura 73
Canal de acetilcolina. A. Estado cerrado. B. DespuS de la unin de la acetilcolina (Ach) y de que un cambio conformacional haya abierto el canal, permitiendo que los iones sodio entren en la fibra muscular y exciten la contraccin. Obsrvense las cargas negati vas en la embocadura del canal, que impiden el paso de iones negativos, como los iones cloruro.
88
UnidadJI
Fisiologa de la membrana, el nervio y el msculo
formacin y liberacin de acetilcolina en esta unin se pro ducen en las siguientes fases: 1. Se forman vesculas pequeas, de aproximadamente 40 nm de tamao, en el aparato de Golgi del cuerpo celular de la motoneurona de la mdula espinal. Estas vesculas son transportadas despus por el axoplasma que fluyea travs del ncleo del axn desde el cuerpo celular central en la mdula espinal hasta la unin neuromuscular en las terminaciones de las fibras nerviosas perifricas. Se acumulan aproximadamente 300.000 de estas pequeas vesculas en las terminaciones nerviosas de una nica placa terminal del msculo esqueltico. 2. La acerilcolina se sintetiza en el citosol de la terminacin de la fibra nerviosa, aunque se transporta inmediatamente a travs de la membrana de las vesculas hasta su interior, donde se almacena en una forma muy concentrada, aproximadamente 10.000 molculas de acetilcolina en cada vescula. 3. Cuando un potencial de accin llega a la terminacin nerviosa. abre muchos canales de calcio en la membrana de la terminacin nerviosa porque esta terminacin tiene muchos canales de calcio activados por el voltaje. En consecuencia, la concentracin de iones calcio en el interior de la membrana terminal aumenta aproximadamente 100 veces.lo que a su vez aumenta la velocidad de fusin de las vesculasde acetilcolina con la membrana terminal aproximadamente 10.000veces. Esta fusin hace que muchas de las vesculas se rompan. permitiendo la exocitosis de la acetilcolina hacia el espacio sinptico. Con cada potencial de accin habitualmente se produce la lisisde aproximadamente 125 vesculas.Posteriormente, despus de algunos milisegundos,la acetilcolina es escindida por la acetilcolinesterasa en ion de acetato y colina, y la colina se reabsorbe activamente en la terminacin neural para ser reutilizada para formar de nuevo acetilcolina. Esta secuencia de acontecimientos se produce en un perodo de 5 a 10milisegundos. 4. El nmero de vesculas disponibles en la terminacin nerviosa es suficiente para permitir la transmisin de slo algunos miles de impulsos desde el nervio hacia el msculo.Por tanto, para la funcin continuada de la unin neuromuscular se deben volver a formar rpidamente nuevas vesculas.En un plazo de algunos segundos,despus de que haya terminado cada uno de los potenciales de accin aparecen hendiduras revestidas en la membrana de la terminacin nerviosa, producidas por las protenas contrctiles de la terminacin nerviosa,especialmente la protena clatrina, que est unida a la membrana en las zonas de las vesculas originales.En un plazo de aproximadamente 20 segundos las protenas se contraen y hacen que las hendiduras se rompan hacia el interior de la membrana, formando de esta manera nuevas vesculas.En un plazo de algunos segundos la acctilcolina es transportada hacia el interior de estas vesculas y ya estn dispuestas para un nuevo ciclo de liberacin de acetilcolina.
+60 +40 +20
ti)
~ ~
(5
O -20 -40 -60 -80 -100 O 15 30 45 Milisegundos 60
:E
75
Potenciales de la placa terminal(en milivoltios). A. Potencial de la placa terminaldebilitado registrado en un msculo curarlzado, de masiado dbil como para generar un potencial de accin. B. Po tencial normal de la placa terminalque desencadena un potencial de accin muscular. C. Potencial de la placa terminal debilitado producido por la toxina botulnica. que reduce la liberacin de acetilcollna en la placa terminal, y que de nuevo es demasiado dbil como para generar un potencial de accin muscular.
tencial de accin. La debilidad del potencial de la placa ter minal del punto A estaba producida por intoxicacin de la fibra muscular con curare, un frmaco que bloquea la ac cin activadora de la acetilcolina sobre los canales de acc tilcolina compitiendo con los puntos del receptor de acetil colina. La debilidad del potencial de la placa terminal del punto se debi al efecto de la toxina botulnica, un vene no bacteriano que reduce la magnitud de la liberacin de acetilcolina por las terminaciones nerviosas.
Factor de seguridad para la transmisin en la unin neuro muscular; fatiga de la unin. Habitualmente cada impulso
que llega a la unin neurornuscular produce un potencial de la placa terminal aproximadamente tres veces mayor gue el necesario para estimular la fibra nerviosa. Por tan to, se dice que la unin neuromuscular normal tiene un elevado factor de seguridad. Sin embargo, la estimulacin de la fibra nerviosa a frecuencias mayores de 100 veces por segundo durante varios minutos con frecuencia dis minuye tanto el nmero de vesculas de acetilcolina que los impulsos no pueden pasar hacia la fibra nerviosa. Esto se denomina fatiga de la unin neurornuscular y es el mis mo efecto que produce fatiga de las sinapsis en el sistema nervioso central cuando las sinapsis son sobreexcitadas. En condiciones normales de funcionamiento raras veces se produce una fatiga medible de la unin neuromuscu lar, e incluso en estos casos slo se ve con los niveles ms intensos de actividad muscular.
Biologa molecular de la formacin y liberacin de acetilcolina
Como la unin neuromuscular es lo suficientemente.grande como para poderla estudiar con facilidad, es una de las po cas sinapsis del sistema nervioso en la que se han estudiado la mayor parte de los detalles de la transmisin qumica. La
Frmacos que potencian o bloquean la transmisin en la unin neuromuscular
Frmacos que estimulan la fibra muscular por su accin simio lar a la acetilcolina. Muchos compuestos, por ejemplo me tacolina, carbaco! y nicotina. tienen el mismo efecto sobre la fibra muscular que la acetilcolina. La diferencia entre es tos frmacos y la acetilcolina consiste en que los frmacos
Captulo 7
Excitacin del msculo esqueltico: transmlsin neuromuscular y acoplamiento excitacin-contraccn
89
no son destruidos por la colinesterasa, () son destruidos tan lentamente que su accin con frecuencia persiste durante muchos minutos a varias horas. Estos frmacos actan pro duciendo zonas localizadas de despolarizacin de la mem brana de la fibra muscular en la placa motora terminal don de estn localizados los receptores de acetilcolina. Despus. cada ver que la fibra muscular se recupera de una contrac cin previa. estas zonas polarizadas, por la fuga de iones. inician un nuevo potencial de accin. produciendo de esta manera un estado de espasmo muscular.
Potencial de accin muscular
Casi todo lo que se ha analizado en el captulo 5 sobre el inicio y la conduccin de los potenciales de accin en las fi bras nerviosas se aplica por igual a las fibras musculares esquelticas, excepto por diferencias cuantitativas. Algu nos de los aspectos cuantitativos de los potenciales muscu lares son los siguientes: 1. Potencial de membrana en reposo: aproximadamente -80 a -90 mV en las fibras esquelticas, el mismo que en las fibras nerviosas rnielinizadas grandes. 2. Duracin del potencial de accin: 1 a 5 ros en el msculo esqueltico, aproximadamente cinco veces mayor que en los nervios mielinizados grandes. 3. Velocidad de conduccin: 3 a 5 mis. aproximadamente 1/13 de la velocidad de conduccin de las fibras nerviosas miclinizadas grandes que excitan al msculo esqueltico.
Frmacos que estimulan la unin neuromuscular mediante la inactivacin de la acetilcolinesterasa. Tres frmacos particu larmente bien conocidos. neostigmina.fisostigmina y fiuoro fosfato de diisopropiio. inactivan la acctilcolinesterasa de las
sinapsis de modo que ya no pueda hidrolizar la acetilcolina. Por tanto. con cada impulso nervioso sucesivo se acumula una cantidad adicional de acetilcolina, que estimula repetitiva mente la fibra muscular. Esto produce espasmomuscularincluso cuando llegan al msculo slo unos pocos impulsos nerviosos. Lamentablemente. tambin puede producir la muerte por es pasmo larngeo. que produce la asfixia del paciente. Neostigmina y fisostigmina se combinan con la acetilco linesterasa para inactivar la acetilcolincsterasa durante has ta varias horas. despus de lo cual estos frmacos son des plazados de la acetilcolinesterasa, de modo que la esterasa es activa de nuevo. Por el contrario, el uorofosfato de diiso propilo, que tiene potencial militar como un potente txico ga seoso nervioso. inactiva la acetilcolinestcrasa durante se manas. lo que hace que sea un txico particularmente letal.
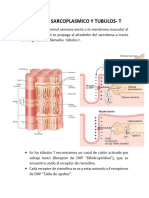
Propagacin del potencial de accin al interior de la fibra muscular a travs de los tbulos transversos
La fibra muscular esqueltica es tan grande que los poten ciales de accin que se propagan a lo largo de la mem brana de su superficie casi no producen ningn flujo de corriente en la profundidad de la fibra. Sin embargo, para producir una contraccin muscular mxima la corriente debe penetrar en las zonas profundas de la fibra muscular hasta la vecindad de las miofibrillas individuales. Esto se consigue mediante la transmisin de los potenciales de ac cin a lo largo de los tbulos transversos (tbulosT), que pe netran a lo largo de toda la fibra muscular desde un extre mo de la fibra hasta el otro, como se seala en la figura 7-5. Los potenciales de accin de los tbulos T producen libe racin de iones calcio en el interior de la fibra muscular en la vecindad inmediata de las miofibrillas, y estos iones cal cio a su vez producen la contraccin. Este proceso global se denomina acoplamiento excitacin-contraccin.
Frmacos que bloquean la transmIsin en la unin "euro muscular. Un grupo de frmacos conocido como frmacos curariformes puede impedir el paso de los impulsos des de la terminacin nerviosa hacia el msculo. Por ejemplo, la n-tubocurarina bloquea la accin de la acetilcolina sobre
los receptores de acetilcolina de la fibra muscular. impi diendo de esta manera el aumento suficiente de la permea bilidad de los canales de la membrana muscular para iniciar un potencial de accin.
Miastenia grave
La miastenia grave. que aparece en aproximadamente 1 de cada 20.000 personas, produce parlisis muscular debido a que las uniones neuromusculares no pueden transmitir sufi cientes seales desde las fibras nerviosas a las fibras muscu lares. En cuanto a su patogenia. en la sangre de la mayor parte de los pacientes que tienen miastcnia grave se han detectado anticuerpos dirigidos frente a las protenas de transporte de los iones sodio activadas por la acetilcolina. Por tanto, se piensa que la miastenia grave es una enferme dad autoinmunitaria en la que los pacientes presentan una respuesta inmunitaria frente a sus propios canales inicos activados por la acetilcolina. Independientemente de la causa. los potenciales de la placa terminal que se producen en las fibras musculares en su mayora son demasiado dbiles para estimular las fibras musculares. Si la enfermedad es lo suficientemente intensa el paciente mucre por parlisis, en particular parlisis de los msculos respiratorios. La enfermedad habitualmente se puede mejorar durante horas mediante la administracin de neostigmino o de cualquier otro fnnaco anticolinestersico. que permite que se acumulen cantidades de acetilcolina ma yores de lo normal en el espacio sinptico. En un plazo de minutos algunas de estas personas paralizadas pueden co menzar a tener una funcin casi normal, basta que sea nece saria una nueva dosis de neostigmina varias horas despus.
Acoplamiento excitacin-contraccin
Sistema de tbulos transversos-retculo sarcoplsmico
La figura 7-5 muestra las miofibrillas rodeadas por el sis tema de tbulos T-retculo sarcoplsrnico. Los tbulos T son muy pequeos y siguen un trayecto transversal a las miofibrillas. Comienzan en la membrana celular y pe netran en todo el espesor desde un lado de la fibra mus cular hasta el lado opuesto. No se muestra en la figura el he cho de que estos tbulos se ramifiquen entre ellos de modo que forman planos completos de tbulos T que se entrelazan entre todas las miofibrillas individuales. Ade ms. donde los tbulos T se originan en la membrana celu lar. estn abiertos hacia el exterior de la fibra muscular. Por tanto, se comunican con el lquido extracelular que rodea la fibra muscular, y ellos mismos contienen lquido extra celular en su luz. En otras palabras. los tbulos T son real mente extensiones internas de la membrana celular. Por
90
Unidad11 Fisiologa de la membrana, el nervio y el msculo
Cisternas terminales
Tbulo transverso
Figura7-5
Sistema tbulo transverso {T)-retfcu lo sarcoplsmico. Obsrvese que los tbulos T se comunican con el exterior de la membrana celular. y que en la profundidad de la fibra muscular cada uno de los tbulosT es adyacente a los extremos de los tbulos longitudinales del retlculo sarcoplsmico que rodean todos los lados de las miofibrillasque en realidad se contraen. Esta ilustra cin se obtuvo de msculo de rana. que tiene un tbulo T por cada sar cmero, localizado en la lnea Z. Se encuentra una disposicin similar en el msculo cardaco de mamfe ros, aunque el msculo esqueltico de mamferos tiene dos tbulos T por cada sarcmero, localizados en las uniones entre las bandas A e 1. (Reproducido a partir de BloomW, Fawcett OW: A Textbook of Histo logy, Philadelphia, WB Saunders, 1986. Modificadode Peachey LO: J Cell Biol 25:209, 1965. Ilustrado por SylviaColard Keene.)
*,#-";---Retrculo sarcoplsmico
Sarcotbulos
tanto, cuando un potencial de accin se propaga por la membrana de una fibra muscular, tambin se propaga un cambio de potencial a lo largo de los tbulos T hacia las zonas profundas del interior de la fibra muscular. De esta manera las corrientes elctricas que rodean a estos tbu los T producen la contraccin muscular. La figura 7-5 tambin muestra un retculo sarcoplsmi co. en amarillo. Est formado por dos partes principales: 1) grandes cavidades denominadas cisternas terminales, que estn junto a los tbulos T, y 2) tbulos longitudinales largos que rodean todas las superficies de las miofibrillas que se estn contrayendo.
branas de las cisternas y tambin en los tbulos longitudi nales unidos a ellas. Estos canales permanecen abiertos durante algunos milisegundos; durante este tiempo se li bera un nmero suficiente de iones calcio hacia el sarco plasma que rodea las miofibrillas para producir la con traccin, como se analiza en el captulo 6.
Bomba de calcio para retirar los iones calcio del lquido mloflbrilar despus de que se haya producido la contrac cin. Una vez que se han liberado los iones calcio desde
los tbulos sarcoplsmicos y que han difundido entre las miofibrillas, la contraccin muscular contina mientras los iones calcio permanezcan a una concentracin eleva da. Sin embargo, una bomba de calcio que acta continua mente y que est localizada en las paredes del retculo sar coplsmico bombea iones calcio desde las miofibrillas de nuevo hacia los tbulos sarcoplsmicos. Esta bomba puede concen trar los iones calcio aproxi madamente 10.000 veces en el interior de los tbulos. Adems, en el interior del re tculo hay una protena denominada calsecuestrina, que puede unirse a hasta 40 veces ms calcio.
Liberacin de iones calcio por el retculo sarcoplsmico
Una de las caractersticas especiales del retculo sarco plsmico es que en el interior de sus tbulos vesiculares hay un exceso de iones calcio a una concentracin eleva da, y que muchos de estos iones son liberados desde cada una de las vesculas cuando se produce un potencial de accin en el t bulo T adyacente. La figura 7-6 muestra que el potencial de accin del t bulo T genera un flujo de corriente hacia las cisternas del retculo sarcoplsmico en su punto de contacto con el t bulo T. Esto, a su vez, da lugar a la apertura rpida de grandes nmeros de canales de calcio en todas las rnern-
"Pulso .. excitador de los iones calcio. La concentracin
normal en estado de reposo (menos de 10-7 molar) de los iones calcio en el citosol que baa las miofibrillas es de masiado pequea como para producir una contraccin. Por tanto, el complejo troponina-troporniosina mantiene
Captulo 7
Excitacin del msculo esqueltico: transmisin neuromuscular y acoplamiento excitacin-contraccin
91
Acoplamiento excitacin-contrac cin en el msculo, que muestra: 1) un potencial de accin que da lugar a la liberacin de iones cal cio desde el retfculo sarcoplsmi co y, posteriormente, 2) recapta cin de los iones calcio por una bomba de calcio.
Rlamentos de actina
Filamentos de miosina
inhibidos los filamentos de actina y mantiene el estado re lajado del msculo. Por el contrario, la excitacin completa del sistema del tbulo T y del retculo sarcoplsmico da lugar a una liberacin de iones calcio suficiente como para aumen tar la concentracin en el lquido miofibrilar basta un valor tan elevado como 2 x 10~ molar, un aumento de 500 veces, que es aproximadamente 10 veces la con centracin necesaria para producir una contraccin muscular mxima. Inmediatamente despus la bomba de calcio produce de nuevo deplecin de los iones cal cio. La duracin total de este pulso de calcio en la fi bra muscular esqueltica normal dura aproximadamente '1/20de segundo, aunque puede durar varias veces ms en algunas fibras y varias veces menos en otras. (En el msculo cardaco el pulso de calcio dura aproximada mente 1/3 de segundo debido a la larga duracin del po tencial de accin cardaco.) Durante este pulso de calcio se produce la contraccin muscular. Si la contraccin debe mantenerse sin interrup ciones durante intervalos prolongados, una serie continua de potenciales de accin repelidos debe iniciar una serie de pulsos de calcio, como se analiza en el captulo 6.
Bibliografa
Vase tambin la bibliografa de los captulos 5 y 6. Allman BL, Rice Cl.: j euromuscular fatigue and aging: central and peripheral facrors. Muscle Nerve 25:785, 2002. Arnonof MJ: Elccrrornyography in Clinical Practice. New York: Churchill Livingstone, 1998. Brown RH Jr: Dysrrophin-associated proteins and the muscular dys trophies. Annu Rev Med 48:457.1997. Chaudhuri A, Behan PO: Fatigue in neurological disorders. Lancer 363:978.2004.
Engel AG, Olmo K Shen XM, Sine SM: Congenital myasrhenic syn dromes: multiple molecular targets at the ncuromuscular junction. Aun NY Acad Sci 998:138,2003. Haouzi P, Chcnuel B, Huszczuk A: Sensing vascular distensin in skeletal muscle by slow conducting afferent fibers: neurophysio logical basis and implication for rcspiratory control. J Appl Phy siol 96:407, 2004. Hoch W: Molecular dissection ofneuromuscular junction formation. Trends Neurosci 26:335, 2003. Keesey JC: Clinical evaluation and rnanagement of myasthenia gravis, Muscle Nerve 29:484,2004. Lee Conformation. acrion, and mechanism ofaction ofneuromus eular blocking muscle relaxants. Pharmacol Ther 98: 143.2003. Leite IF, Rodrigues-Pmguet N, Lester HA: Tnsights into channel function via channel dysfunction. J Clin lnvest III :436,2003. Payne AM, Dclbono O: Ncurogenesis of excitarion-conrraction un eoupling in aging skeletal muscle. Exerc Sport Sci Rev 32:36, 2004. Pene D: Historical perspcctives: plasticity of rnammalian skeJetal muscle. J Appl Physiol 90: 11 19. 2001. Rekling lC, Funk GD, Bayliss DA, et al: Synaptic control ofmoto neuronal excitability, Physiol Rev 80:767. 2000. Schiaffino S, Serrano A: Calcineurin signaling and neural control of skeetal muscle fiber rypc ami size. Trcnds Pbarmacol Sci 23:569, 2002. Tang W, Sencer S, Hamilton SI..: Calmodulin modulation of proteins involved in excitation-comraction coupling. From Biosci 7:583. 2002. Toyoshima C, ornura H. Sugita Y: Strucrural basis of ion pumping by eal+-ATPase of sarcoplasmic reticulum. fEBS Leu 555: 106, 2003. Van der Kloot W. vlolgo J: Quantal acerylcholine release al the ver tebrate neuromuscular junction. Physiol Rev 74:899, 1994. VincentA: Unraveling the pathogenesis of myasthenia gravis. NatRev lmmunol 10:797,2002. Vincent A, McConville J, Farrugia ME, er al: Antibodies in myast henia gravis and relatcd disorders. Ann N Y Acad Sci 998:324. 2003.
e:
Potrebbero piacerti anche
- Desgrabaciones. Farmacología II. Vasodilatadores: Dra. Ysrael Centeno Semestre I-2023Documento29 pagineDesgrabaciones. Farmacología II. Vasodilatadores: Dra. Ysrael Centeno Semestre I-2023fabrizioNessuna valutazione finora
- C9 FisioDocumento16 pagineC9 FisioBenjamin Vega MNessuna valutazione finora
- Musculo Liso Celulas ContractilesDocumento64 pagineMusculo Liso Celulas ContractilesmhuancasNessuna valutazione finora
- Práctica 21 - Grupo7Documento10 paginePráctica 21 - Grupo7RIVALDO ALFREDO RIVEROS POLONessuna valutazione finora
- Capitulo 7Documento6 pagineCapitulo 7MisaelNessuna valutazione finora
- La Excitación y Contracción Del Músculo LisoDocumento5 pagineLa Excitación y Contracción Del Músculo LisoBianca GonzalezNessuna valutazione finora
- Potencial de Accion Corazón y FármacosDocumento10 paginePotencial de Accion Corazón y FármacosmcordovaromNessuna valutazione finora
- Apuntes Biofisica Musculo CardiacoDocumento33 pagineApuntes Biofisica Musculo CardiacoCamila NievesNessuna valutazione finora
- Fisio FinalDocumento256 pagineFisio FinalRicardo CarrilloNessuna valutazione finora
- 7 Fármacos Activadores de Receptores Colinérgicos e Inhibidores de ColinesterasaDocumento11 pagine7 Fármacos Activadores de Receptores Colinérgicos e Inhibidores de ColinesterasaArne SaenzNessuna valutazione finora
- Transmision NeuromuscularDocumento6 pagineTransmision NeuromuscularHoracio Ramìrez LopezNessuna valutazione finora
- Control Nervioso y Hormonal Del Músculo LisoDocumento14 pagineControl Nervioso y Hormonal Del Músculo LisoMichell Vidal AnsNessuna valutazione finora
- Guia 4 Contraccion Muscular 2021-1Documento17 pagineGuia 4 Contraccion Muscular 2021-1HOLMAN AUDREY ALBA CASTRONessuna valutazione finora
- CAPÍTULO 21 Obstetricia de Williams - Fisiología Del Trabajo de PartoDocumento33 pagineCAPÍTULO 21 Obstetricia de Williams - Fisiología Del Trabajo de PartoNataly TobarNessuna valutazione finora
- Haga Un Diagrama Del Metabolismo de Los NeurotransmisoresDocumento6 pagineHaga Un Diagrama Del Metabolismo de Los NeurotransmisoresCarlos Jair Herrera CamposNessuna valutazione finora
- Ordi Fisio I ResumenesDocumento70 pagineOrdi Fisio I ResumenesLupita PerezNessuna valutazione finora
- Cap. 4 y 5Documento45 pagineCap. 4 y 5Genesis RiveroNessuna valutazione finora
- Curares 2017Documento27 pagineCurares 2017anon_449996588Nessuna valutazione finora
- Excitacion y Contraccion Del Musculo LisoDocumento40 pagineExcitacion y Contraccion Del Musculo LisoAndrea MuñozNessuna valutazione finora
- Musculo Esqueletico, Liso y Cardiaco.Documento10 pagineMusculo Esqueletico, Liso y Cardiaco.Veronica LopezNessuna valutazione finora
- Canales Ionicos Regulados Por VoltajeDocumento24 pagineCanales Ionicos Regulados Por VoltajeAlex Madera100% (1)
- Farmacología de Los Simpaticomiméticos Indicaciones TerapéuticasDocumento16 pagineFarmacología de Los Simpaticomiméticos Indicaciones TerapéuticasYordanNessuna valutazione finora
- Bloqueadores de Los Canales de CalcioDocumento55 pagineBloqueadores de Los Canales de Calciodayana esparragozaNessuna valutazione finora
- Informe de Sesión 2Documento8 pagineInforme de Sesión 2SHIRLEY OLENKA HONORIO BRICEÑONessuna valutazione finora
- 7.contracción Muscular y Tipo de MúsculosDocumento9 pagine7.contracción Muscular y Tipo de MúsculosMelany RodríguezNessuna valutazione finora
- Práctica 7 Potencial de Accion Corazón y Fármacos Con DatosDocumento16 paginePráctica 7 Potencial de Accion Corazón y Fármacos Con Datosyefferson silva sullonNessuna valutazione finora
- Clase 4 FisiologíaDocumento10 pagineClase 4 FisiologíaIsrael Peña RivasNessuna valutazione finora
- TPmusculolisoconrtas 2023Documento3 pagineTPmusculolisoconrtas 2023BarbaraNessuna valutazione finora
- Repaso Tema 2Documento6 pagineRepaso Tema 2Sebastian GonzalezNessuna valutazione finora
- Clase 3-Potencial de AcciónDocumento14 pagineClase 3-Potencial de AcciónROMÁN ELEAZAR OJEDA ZAPATANessuna valutazione finora