Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- 6th Central Pay Commission Salary CalculatorDocumento15 pagine6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Islamic Accounting-Article 3Documento7 pagineIslamic Accounting-Article 3Mohd Firdaus Abd LatifNessuna valutazione finora
- Independent and Dependent Variables ScenariosDocumento2 pagineIndependent and Dependent Variables ScenariosPaula Krissha Ramos50% (2)
- Oil EOR HandbookDocumento24 pagineOil EOR HandbookRicardo Hernan Gomez BNessuna valutazione finora
- Media Role DemocraticDocumento22 pagineMedia Role DemocraticMohd Firdaus Abd LatifNessuna valutazione finora
- Interim Income Tax Accounting 070919 SigninsheetDocumento3 pagineInterim Income Tax Accounting 070919 SigninsheetMohd Firdaus Abd LatifNessuna valutazione finora
- Cpaverify ReportDocumento1 paginaCpaverify ReportMohd Firdaus Abd LatifNessuna valutazione finora
- AES (Automated Defense System)Documento6 pagineAES (Automated Defense System)Mohd Firdaus Abd LatifNessuna valutazione finora
- New Media The New Economy and New SpacesDocumento35 pagineNew Media The New Economy and New SpacesMohd Firdaus Abd LatifNessuna valutazione finora
- BMD401 Mandarin ScriptDocumento3 pagineBMD401 Mandarin ScriptMohd Firdaus Abd LatifNessuna valutazione finora
- Media Prima ANNUAL REPORT 2011Documento248 pagineMedia Prima ANNUAL REPORT 2011Mohd Firdaus Abd LatifNessuna valutazione finora
- Men in PRDocumento8 pagineMen in PRMohd Firdaus Abd LatifNessuna valutazione finora
- The Research ProposalDocumento7 pagineThe Research ProposalShubhangi HingoleNessuna valutazione finora
- Law of HududDocumento8 pagineLaw of HududMohd Firdaus Abd LatifNessuna valutazione finora
- Media Law HandbookDocumento74 pagineMedia Law Handbookuppercaise100% (1)
- Media Law HandbookDocumento74 pagineMedia Law Handbookuppercaise100% (1)
- Electronic Mass Media LawDocumento36 pagineElectronic Mass Media LawMohd Firdaus Abd LatifNessuna valutazione finora
- Term PaperDocumento10 pagineTerm PaperMohd Firdaus Abd LatifNessuna valutazione finora
- Comparison of Hydrolysis Conditions To Recover Reducing Sugar From Various Lignocellulosic MaterialsDocumento11 pagineComparison of Hydrolysis Conditions To Recover Reducing Sugar From Various Lignocellulosic MaterialsMohd Firdaus Abd LatifNessuna valutazione finora
- Guide To Mini ResearchDocumento3 pagineGuide To Mini ResearchMohd Firdaus Abd LatifNessuna valutazione finora
- Resume Muhammad Faridzul Bin Zainal AbidinDocumento7 pagineResume Muhammad Faridzul Bin Zainal AbidinMohd Firdaus Abd LatifNessuna valutazione finora
- Research Topic IdeaDocumento5 pagineResearch Topic IdeaMohd Firdaus Abd LatifNessuna valutazione finora
- Lecture On Mass Media SocietyDocumento70 pagineLecture On Mass Media SocietyMohd Firdaus Abd LatifNessuna valutazione finora
- Media Plan For Harley DaviddsonDocumento43 pagineMedia Plan For Harley DaviddsonMohd Firdaus Abd LatifNessuna valutazione finora
- Study Guide Cells Test AnswersDocumento10 pagineStudy Guide Cells Test Answersapi-267855902100% (1)
- Summative Assessment QuizDocumento3 pagineSummative Assessment Quizapi-379149339Nessuna valutazione finora
- SH Libro CompletoDocumento80 pagineSH Libro CompletoMickel Caballero100% (1)
- Drug Development ProcessDocumento16 pagineDrug Development ProcessNachiket PatelNessuna valutazione finora
- Why We Need To Protect Polar BearsDocumento6 pagineWhy We Need To Protect Polar BearsTrân ĐinhNessuna valutazione finora
- Treatment of Latent Tuberculosis InfectionDocumento10 pagineTreatment of Latent Tuberculosis InfectionRichard WooliteNessuna valutazione finora
- Contents of Practicals XIIDocumento3 pagineContents of Practicals XIINibedeetaNessuna valutazione finora
- Lucienne NeoMASS AAAC PlusDocumento8 pagineLucienne NeoMASS AAAC PlusRanjith EswaranNessuna valutazione finora
- Parameter Estimation With Term-Wise Decomposition in Biochemical Network GMA Models by Hybrid Regularized Least Squares-Particle Swarm OptimizationDocumento8 pagineParameter Estimation With Term-Wise Decomposition in Biochemical Network GMA Models by Hybrid Regularized Least Squares-Particle Swarm OptimizationGek CagatanNessuna valutazione finora
- Arndt Schmidt-Denter Auer Weitere 2003Documento15 pagineArndt Schmidt-Denter Auer Weitere 2003Louis Fetilo FabunanNessuna valutazione finora
- Silviculture: February 2018Documento69 pagineSilviculture: February 2018Tang YinNessuna valutazione finora
- (English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)Documento78 pagine(English (Auto-Generated) ) Ep. 94 - Andres Ryan of BioAquatix - Com On Breeding Clown Loaches (DownSub - Com)acromontiNessuna valutazione finora
- Orientation in Paraclinic ScienceDocumento13 pagineOrientation in Paraclinic ScienceShivani YadavNessuna valutazione finora
- Lecture 3 Cell Cycle - QuestionsDocumento6 pagineLecture 3 Cell Cycle - Questions中华雅思王Nessuna valutazione finora
- Al-Zahraa Interchange: Section A-ADocumento1 paginaAl-Zahraa Interchange: Section A-ANadher Al-BaghdadiNessuna valutazione finora
- Imperata Cylindrica: Investigations On Some Aspects of Chemical Ecology of Cogongrass, (L.) BeauvDocumento10 pagineImperata Cylindrica: Investigations On Some Aspects of Chemical Ecology of Cogongrass, (L.) BeauvDiroy ParulianNessuna valutazione finora
- MCQ of Diagnosis BacterialDocumento11 pagineMCQ of Diagnosis Bacterialمحمد اللاميNessuna valutazione finora
- Physiology vs. Anatomy: by Sarah C.P. Willaims Medically Reviewed by Minesh Khatri, MD On July 28, 2021Documento2 paginePhysiology vs. Anatomy: by Sarah C.P. Willaims Medically Reviewed by Minesh Khatri, MD On July 28, 2021exo oxeNessuna valutazione finora
- PK PD en Critical IllnessDocumento27 paginePK PD en Critical IllnessDaniela BolivarNessuna valutazione finora
- MotivationDocumento2 pagineMotivationPrachi Srivastava100% (1)
- Effect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly PopulationDocumento6 pagineEffect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly Populationastrinila fauziNessuna valutazione finora
- Q1 Module 3 Science 9Documento32 pagineQ1 Module 3 Science 9リリスセレインNessuna valutazione finora
- HINANAY-HENIELOUISE-D - Activity 1 (Intro - in Microbiology and Parasitology)Documento6 pagineHINANAY-HENIELOUISE-D - Activity 1 (Intro - in Microbiology and Parasitology)Henie Louise HinanayNessuna valutazione finora
- Wed09 NovozymesDocumento29 pagineWed09 NovozymesPulbere NeagraNessuna valutazione finora
- Quiz Bowl Competition 2019: Theme: MetabolizeDocumento77 pagineQuiz Bowl Competition 2019: Theme: MetabolizeKazuya SenseiNessuna valutazione finora
- Statement Uni - SenithaDocumento2 pagineStatement Uni - SenithaSenitha MindulaNessuna valutazione finora
- BAM Chapter 29. CronobacterDocumento14 pagineBAM Chapter 29. CronobacterremyNessuna valutazione finora
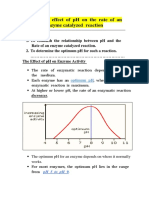
- Effect of PH On Enzyme ActivityDocumento12 pagineEffect of PH On Enzyme ActivityAb AbNessuna valutazione finora