Potrebbero piacerti anche
- Theories and Constructs of RaceDocumento16 pagineTheories and Constructs of RaceX69hotdog100% (1)
- Color Environment and Human Response PDFDocumento2 pagineColor Environment and Human Response PDFMario100% (1)
- What's New - ChangelogDocumento6 pagineWhat's New - ChangelogErnesto De LeonNessuna valutazione finora
- Albinism Classification, Clinical.23Documento4 pagineAlbinism Classification, Clinical.23Danny WongNessuna valutazione finora
- Structure and Functions of HaemoglobinDocumento2 pagineStructure and Functions of HaemoglobinKrissy KayNessuna valutazione finora
- Biology N5 Past Paper Questions Cell Biology1Documento46 pagineBiology N5 Past Paper Questions Cell Biology1moizNessuna valutazione finora
- Chapter 1 - Science of Zoology NotesDocumento7 pagineChapter 1 - Science of Zoology Notesapi-195601294Nessuna valutazione finora
- Extraction, Isolation, Purification and Identification CurcuminDocumento16 pagineExtraction, Isolation, Purification and Identification CurcuminRaisyIkrimahNessuna valutazione finora
- Population Genetics ProblemsDocumento4 paginePopulation Genetics ProblemsRahul KumarNessuna valutazione finora
- Bond & Cash JASP 1992Documento15 pagineBond & Cash JASP 1992Jameel Ricardo HugginsNessuna valutazione finora
- Reading Wuiz Hazırlık RE3 1-3Documento13 pagineReading Wuiz Hazırlık RE3 1-3Kübra Şendoğan100% (1)
- Race Without Color PDFDocumento5 pagineRace Without Color PDFAmoklerougeNessuna valutazione finora
- Chromabright Skin Brightener PresentationDocumento22 pagineChromabright Skin Brightener PresentationRnD Roi SuryaNessuna valutazione finora
- Signature Assignment Paper Anth 2Documento8 pagineSignature Assignment Paper Anth 2api-301347675Nessuna valutazione finora
- Mind Your Face ShapeDocumento52 pagineMind Your Face ShapeYana Zinovyeva100% (3)
- AlbinismDocumento3 pagineAlbinismNaim ElmasriNessuna valutazione finora
- Chromosomal Abnormalities in Humans and AnimalsDocumento15 pagineChromosomal Abnormalities in Humans and AnimalsGautam ChandwaniNessuna valutazione finora
- AMYLOIDOSIS: Lecture Materials For StudentsDocumento134 pagineAMYLOIDOSIS: Lecture Materials For StudentsgechanatomyNessuna valutazione finora
- Polytene and Lampbrush ChromosomesDocumento3 paginePolytene and Lampbrush Chromosomesneeru.bhagatNessuna valutazione finora
- MTT AssayDocumento2 pagineMTT AssayHameedhaNessuna valutazione finora
- Harmful Effects of Gamma RaysDocumento3 pagineHarmful Effects of Gamma RaysKresha ParreñasNessuna valutazione finora
- What Is Pompe Disease and What Are The Causes of Pompe Disease?Documento4 pagineWhat Is Pompe Disease and What Are The Causes of Pompe Disease?Gamze Hatlar100% (1)
- Chromosomal AbnormalitiesDocumento3 pagineChromosomal AbnormalitiesYra Mie DitaroNessuna valutazione finora
- IndometacinDocumento47 pagineIndometacinSava1988Nessuna valutazione finora
- Arms ProtocolDocumento15 pagineArms ProtocolMahmOod Gh0% (1)
- Genetic Disorders: Ma. Minda Luz M. Manuguid, M.DDocumento65 pagineGenetic Disorders: Ma. Minda Luz M. Manuguid, M.Djulo_05100% (1)
- IHC - InterpretareDocumento185 pagineIHC - InterpretareAnca NeaguNessuna valutazione finora
- Principle and Applications of Gel Fitration ChromatographyDocumento18 paginePrinciple and Applications of Gel Fitration ChromatographyGanesh V GaonkarNessuna valutazione finora
- Transfer RNA Synthesis and Regulation: Advanced ArticleDocumento17 pagineTransfer RNA Synthesis and Regulation: Advanced ArticleAfif BerlianNessuna valutazione finora
- ToxicologieDocumento29 pagineToxicologieRazvan FumeaNessuna valutazione finora
- Linkage, Crossing-Over, & GeneDocumento13 pagineLinkage, Crossing-Over, & GeneAbel ClaireNessuna valutazione finora
- Lecture - 1 Hematological StainingDocumento20 pagineLecture - 1 Hematological StainingOjambo FlaviaNessuna valutazione finora
- 3 PP Animal BehaviorDocumento21 pagine3 PP Animal BehaviorMesut DenizNessuna valutazione finora
- M - 23 Polytene ChromosomeDocumento3 pagineM - 23 Polytene ChromosomeDr. Tapan Kr. DuttaNessuna valutazione finora
- Allium Cepa Test 1Documento5 pagineAllium Cepa Test 1Chinkee Illanza LimNessuna valutazione finora
- Numerical Aberrations: Meiotic NondisjunctionDocumento36 pagineNumerical Aberrations: Meiotic NondisjunctionHemanth_Prasad_2860Nessuna valutazione finora
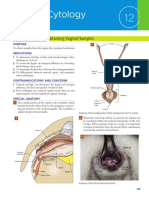
- Vaginal CytologyDocumento4 pagineVaginal Cytologyابراهيم عبدالله100% (1)
- ArginaseDocumento5 pagineArginasecesarjavier1290Nessuna valutazione finora
- Cadmium PollutionDocumento13 pagineCadmium PollutionRima ArisnawatiNessuna valutazione finora
- CellDocumento47 pagineCellAnami NizamNessuna valutazione finora
- 1378968054.1856 - Histochemistry and Enzyme HistochemistryDocumento39 pagine1378968054.1856 - Histochemistry and Enzyme HistochemistryCarlos Ivan Cruz Cardenas100% (1)
- Classification of Genetic DisordersDocumento6 pagineClassification of Genetic DisordersshobhaNessuna valutazione finora
- 1 Cervical and Vaginal CytologyDocumento63 pagine1 Cervical and Vaginal Cytologynanxtoyah100% (1)
- CitologieDocumento14 pagineCitologieAlexandra CrisanNessuna valutazione finora
- HALOPHYTESDocumento8 pagineHALOPHYTESGobiNessuna valutazione finora
- The Normal and The PathologicalDocumento14 pagineThe Normal and The PathologicalnluxonNessuna valutazione finora
- Clinical Features and Molecular DiagnosisDocumento102 pagineClinical Features and Molecular Diagnosiskholoud220Nessuna valutazione finora
- Hematological Genetic Disorders A ReviewDocumento18 pagineHematological Genetic Disorders A ReviewShodh Sangam - A RKDF University Journal of Science and EngineeringNessuna valutazione finora
- Lung Tumor PDFDocumento116 pagineLung Tumor PDFChetan Kumar100% (1)
- ACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFDocumento12 pagineACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFLpNessuna valutazione finora
- The Grignard ReagentDocumento3 pagineThe Grignard ReagentPris97Nessuna valutazione finora
- Protein: Proteins (Documento17 pagineProtein: Proteins (subhasis1172Nessuna valutazione finora
- Lysosomes PresentationDocumento53 pagineLysosomes PresentationCaroline Stephenson50% (2)
- NeisseriaDocumento33 pagineNeisseriaShuler0071Nessuna valutazione finora
- Digestion LabDocumento7 pagineDigestion LabnicewanNessuna valutazione finora
- AlbinismDocumento10 pagineAlbinismTapash GopeNessuna valutazione finora
- Antimicrobial Peptides: From Wikipedia, The Free EncyclopediaDocumento15 pagineAntimicrobial Peptides: From Wikipedia, The Free EncyclopediaAnonymous 679hyCDUNessuna valutazione finora
- IsomersDocumento5 pagineIsomersSobia Saleem100% (1)
- 01 Neoplastic Diseases of The UterusDocumento11 pagine01 Neoplastic Diseases of The Uterusi_top3Nessuna valutazione finora
- Hemoglobin Structure & SynthesisDocumento24 pagineHemoglobin Structure & SynthesisIMDCBiochemNessuna valutazione finora
- HER2 4B5 Interpretation GuideDocumento16 pagineHER2 4B5 Interpretation GuideAndres RestrepoNessuna valutazione finora
- 2 CLab ManualDocumento129 pagine2 CLab ManualMomerNessuna valutazione finora
- TALASEMIADocumento1 paginaTALASEMIAmas bambangNessuna valutazione finora
- Amyloid StainsDocumento7 pagineAmyloid Stainsridhisood16Nessuna valutazione finora
- An Overview of MetabolismDocumento6 pagineAn Overview of MetabolismFieska AzizahNessuna valutazione finora
- Cell Biology Practice 2Documento15 pagineCell Biology Practice 2NgMinhHaiNessuna valutazione finora
- Evidence of Functional Redundancy Between MID Proteins: Implications For The Presentation of Opitz SyndromeDocumento8 pagineEvidence of Functional Redundancy Between MID Proteins: Implications For The Presentation of Opitz SyndromeSolangel Burbano VillotaNessuna valutazione finora
- Pigmented Oral LesionDocumento133 paginePigmented Oral LesionArnela UniqueNessuna valutazione finora
- Race As A ClassDocumento6 pagineRace As A ClassTim DoNessuna valutazione finora
- Ten Things Everyone Should Know About RaceDocumento1 paginaTen Things Everyone Should Know About RacemargaretNessuna valutazione finora
- Evolution of Skin ColorDocumento2 pagineEvolution of Skin ColorMary TaklaNessuna valutazione finora
- Brochure Zeropollution 2019 LowDocumento6 pagineBrochure Zeropollution 2019 LowScarlethMuñozNessuna valutazione finora
- Applications in Cosmetics: Preparation and Application in IndustryDocumento67 pagineApplications in Cosmetics: Preparation and Application in IndustryAchmad LatiefNessuna valutazione finora
- Integumentary SystemDocumento54 pagineIntegumentary SystemKhaela Mercader100% (2)
- Kobo AnOverviewofFoundationMake upforDiverseSkinTones Dec2016Documento6 pagineKobo AnOverviewofFoundationMake upforDiverseSkinTones Dec2016anuyasane9776Nessuna valutazione finora
- Loraine Jane Clamonte Done 3Documento38 pagineLoraine Jane Clamonte Done 3Lorena NavarroNessuna valutazione finora
- 10.2 Student SheetDocumento34 pagine10.2 Student Sheetmike bevnNessuna valutazione finora
- The Complete Guide To Vitiligo Corner 2010Documento107 pagineThe Complete Guide To Vitiligo Corner 2010mohanarameshNessuna valutazione finora
- Byrd, Ceacal, Felton, Nicholson, Rhaney, McCray, Young 2017Documento18 pagineByrd, Ceacal, Felton, Nicholson, Rhaney, McCray, Young 2017ZeynepNessuna valutazione finora
- Dermatologica Sinica: Ai-Young LeeDocumento7 pagineDermatologica Sinica: Ai-Young LeeNatta ApucaranaNessuna valutazione finora
- Jarvis 12 Skin Hair NailsDocumento11 pagineJarvis 12 Skin Hair NailsSarah C. SnooksNessuna valutazione finora
- Addressing The Skin Colour Discrimination in AdsDocumento2 pagineAddressing The Skin Colour Discrimination in AdsMohammad Habibur RahmanNessuna valutazione finora
- A "Lost White Tribe" That Created A Global Racial CasteDocumento330 pagineA "Lost White Tribe" That Created A Global Racial CasteAsar100% (2)
- Skin: The Body's Canvas: External Chemicals LeisureDocumento2 pagineSkin: The Body's Canvas: External Chemicals LeisureSon PhamNessuna valutazione finora
- Etheric MelaninDocumento40 pagineEtheric MelaninAsar100% (1)
- Process Portfolio 1 - MinDocumento26 pagineProcess Portfolio 1 - Minapi-592618881Nessuna valutazione finora
- MZ SKIN Training Manual 2023Documento56 pagineMZ SKIN Training Manual 2023ZB ChuaNessuna valutazione finora
- Plagiarism Scan Report: Plagiarised UniqueDocumento2 paginePlagiarism Scan Report: Plagiarised UniqueAneeqaNessuna valutazione finora