Potrebbero piacerti anche
- Randomly Amplified DNA Fingerprinting: A Culmination of DNA Marker Technologies Based On Arbitrarily-Primed PCR AmplificationDocumento11 pagineRandomly Amplified DNA Fingerprinting: A Culmination of DNA Marker Technologies Based On Arbitrarily-Primed PCR AmplificationIndah Maulidya IriantiNessuna valutazione finora
- RFLP and RapdDocumento13 pagineRFLP and Rapdsaboor saifiNessuna valutazione finora
- Molecular Markers and Their Characteristics M. R. Meena, M.L. Chhabra, Ravinder Kumar and C. M. MeenaDocumento5 pagineMolecular Markers and Their Characteristics M. R. Meena, M.L. Chhabra, Ravinder Kumar and C. M. Meenamintu_uasNessuna valutazione finora
- Safety of Students GuidelinesDocumento18 pagineSafety of Students GuidelinesdrugdrugNessuna valutazione finora
- PCR Based Molecualr, Genetic MarkersDocumento59 paginePCR Based Molecualr, Genetic MarkersRafay MirzaNessuna valutazione finora
- RFLP & RapdDocumento25 pagineRFLP & RapdBalaji Prasanna KumarNessuna valutazione finora
- Molecular Marker:: Quality For A Good Genetic MarkerDocumento5 pagineMolecular Marker:: Quality For A Good Genetic MarkerRakesh GedalaNessuna valutazione finora
- Product Manual: GL RAPD Decamer SetsDocumento35 pagineProduct Manual: GL RAPD Decamer SetsAnonymous C6laqkPAAKNessuna valutazione finora
- Mol Breed - 2Documento20 pagineMol Breed - 2Sukanta RoyNessuna valutazione finora
- NAMEDocumento11 pagineNAMEEmpowering YouthNessuna valutazione finora
- Molecular Markers: Types of Genetic MarkersDocumento9 pagineMolecular Markers: Types of Genetic Markerssunayana debNessuna valutazione finora
- Restriction Enzyme (Restriction Endonuclease)Documento18 pagineRestriction Enzyme (Restriction Endonuclease)yusuf zihniNessuna valutazione finora
- Polymerase Chain Reaction (PCR)Documento67 paginePolymerase Chain Reaction (PCR)Derick SemNessuna valutazione finora
- 4B-Molecular MarkerDocumento67 pagine4B-Molecular MarkerSharvind Kumar Sharvind KumarNessuna valutazione finora
- DNA FingerprintingDocumento83 pagineDNA Fingerprintinghamna farooqiNessuna valutazione finora
- Agarwal 2008 - Advances in Molecular Marker Techniques and Their Applications in Plant SciencesDocumento16 pagineAgarwal 2008 - Advances in Molecular Marker Techniques and Their Applications in Plant SciencessherenelaiNessuna valutazione finora
- Applications of RAPDDocumento9 pagineApplications of RAPDFrancisNessuna valutazione finora
- Concise Notes Summary Genetic Engineering PCRDocumento26 pagineConcise Notes Summary Genetic Engineering PCRVoice AnonymousNessuna valutazione finora
- DNA Polymorphisms Amplified by Arbitrary Primers Are Useful As Genetic Markers PRDocumento9 pagineDNA Polymorphisms Amplified by Arbitrary Primers Are Useful As Genetic Markers PRSheryl AnNessuna valutazione finora
- Thierry Huguet, György B. Kiss, Attila Kereszt, Dong-Jin Kim and Doug CookDocumento31 pagineThierry Huguet, György B. Kiss, Attila Kereszt, Dong-Jin Kim and Doug CookSantosh TummaNessuna valutazione finora
- 2017 - RNAseq - Methods For Transcriptome AnalysisDocumento17 pagine2017 - RNAseq - Methods For Transcriptome AnalysisAntarToumaNessuna valutazione finora
- RFLP, Rapd & AflpDocumento32 pagineRFLP, Rapd & AflpArun Kumar Boddapati100% (1)
- Molecular Markers in Forest Management and Tree Breeding: A ReviewDocumento7 pagineMolecular Markers in Forest Management and Tree Breeding: A ReviewFernando Castro EchavezNessuna valutazione finora
- DNA SequencingDocumento31 pagineDNA Sequencingmymobile502100% (1)
- DNA Based Molecular MarkersDocumento21 pagineDNA Based Molecular MarkersShubham PrasadNessuna valutazione finora
- Amplified Fragment Length Polymorphism (AFLP) : A Review of The Procedure and Its ApplicationsDocumento16 pagineAmplified Fragment Length Polymorphism (AFLP) : A Review of The Procedure and Its ApplicationsDario Farias CantillanaNessuna valutazione finora
- Molecular MarkersDocumento4 pagineMolecular MarkersKai RobinsonNessuna valutazione finora
- Aflp-Pcr:: Steps: 1.DNA ExtractionDocumento3 pagineAflp-Pcr:: Steps: 1.DNA ExtractionArsalan Abbas ZafarNessuna valutazione finora
- Proteomics and GenomicsDocumento16 pagineProteomics and GenomicsPAVITHRA VNessuna valutazione finora
- CLC Genetics Chapter 3Documento51 pagineCLC Genetics Chapter 3Châu Anh TrầnNessuna valutazione finora
- PCRWksheetDocumento2 paginePCRWksheetKevin KocotNessuna valutazione finora
- Molecular MarkersDocumento37 pagineMolecular MarkersJeremiah Christopher ManzanoNessuna valutazione finora
- Lecture 2 - Molecular MarkersDocumento59 pagineLecture 2 - Molecular MarkersDeepali SinghNessuna valutazione finora
- Molecular Markers in Plant Genome AnalysisDocumento13 pagineMolecular Markers in Plant Genome Analysisarunnene2Nessuna valutazione finora
- HistoryDocumento14 pagineHistoryAditi ShresthaNessuna valutazione finora
- Molecular MarkerDocumento26 pagineMolecular MarkershivamNessuna valutazione finora
- Molecular Marker: Study NotesDocumento14 pagineMolecular Marker: Study NotesSasikala RajendranNessuna valutazione finora
- Development of A Rapid and Inexpensive Method To Reveal Natural Antisense TranscriptsDocumento8 pagineDevelopment of A Rapid and Inexpensive Method To Reveal Natural Antisense Transcriptschandran.vkrsdNessuna valutazione finora
- Inter-Simple Sequence Repeat (ISSR) MarkersDocumento11 pagineInter-Simple Sequence Repeat (ISSR) MarkersMihaela MihaNessuna valutazione finora
- Northern Blot WikiDocumento5 pagineNorthern Blot WikiTeuku IndraNessuna valutazione finora
- Definition of Genetic MarkersDocumento16 pagineDefinition of Genetic MarkersAisha Warshaf100% (1)
- Continuous Assessment: Concentration's Increment in Case of Insert and LigaseDocumento8 pagineContinuous Assessment: Concentration's Increment in Case of Insert and LigaseDebapriya HazraNessuna valutazione finora
- V08 - Yvan Le Bras - Training RADSeq - 0 PDFDocumento55 pagineV08 - Yvan Le Bras - Training RADSeq - 0 PDFSuany Quesada CalderonNessuna valutazione finora
- Brugmans Et Al. 2002 (Transcript Mapping)Documento12 pagineBrugmans Et Al. 2002 (Transcript Mapping)Sathish KumarNessuna valutazione finora
- Introduction To Molecular MarkersDocumento47 pagineIntroduction To Molecular MarkersSama MouradNessuna valutazione finora
- Genetic Engineering and Recombinant DNA Technology: Ashok Kumar (70700012) Ila Chawla (70700019)Documento54 pagineGenetic Engineering and Recombinant DNA Technology: Ashok Kumar (70700012) Ila Chawla (70700019)Harshit KumarNessuna valutazione finora
- Chapter 13 - Molecular MethodsDocumento29 pagineChapter 13 - Molecular MethodsYoiceMartinaPawekaNessuna valutazione finora
- In Situ Hybridization: Preparation of MaterialDocumento12 pagineIn Situ Hybridization: Preparation of MaterialchablainNessuna valutazione finora
- Mol Breed - 1 - 7 - MergedDocumento88 pagineMol Breed - 1 - 7 - MergedSukanta RoyNessuna valutazione finora
- Mol Breed - 1Documento21 pagineMol Breed - 1Sukanta RoyNessuna valutazione finora
- DNA Based Techniques For Studying Genetic DiversityDocumento30 pagineDNA Based Techniques For Studying Genetic DiversitySyahrul Tuba Al FatihNessuna valutazione finora
- Punam Jaiswal Pg-III Dna FingerprintingDocumento7 paginePunam Jaiswal Pg-III Dna FingerprintingPrathamesh KharabeNessuna valutazione finora
- DNA Detection Using Recombination Proteins: BiologyDocumento7 pagineDNA Detection Using Recombination Proteins: BiologyclchioNessuna valutazione finora
- Aug. 31 - Sept. 4, 2014Documento49 pagineAug. 31 - Sept. 4, 2014giapvanthuNessuna valutazione finora
- 1516260242FSC P13 M13 E-Text PDFDocumento10 pagine1516260242FSC P13 M13 E-Text PDFKanchanNessuna valutazione finora
- Unit 1 Amplification TechniqueDocumento9 pagineUnit 1 Amplification TechniqueShin BoyceNessuna valutazione finora
- Molecular Biology & R-DNA TechnologyDocumento12 pagineMolecular Biology & R-DNA TechnologyRajnesh ParasharNessuna valutazione finora
- Klindworth 2013 Evaluation of General S Ribosomal RDocumento11 pagineKlindworth 2013 Evaluation of General S Ribosomal RVisa LaserNessuna valutazione finora
- RNA Methodologies: A Laboratory Guide for Isolation and CharacterizationDa EverandRNA Methodologies: A Laboratory Guide for Isolation and CharacterizationNessuna valutazione finora
- Lowry SwitchgrassDocumento11 pagineLowry SwitchgrassDewi Kartika DekaNessuna valutazione finora
- (P) Random Amplified PolymorphicDNA (RAPD) Markers PDFDocumento12 pagine(P) Random Amplified PolymorphicDNA (RAPD) Markers PDFDewi Kartika DekaNessuna valutazione finora
- (P) Random Amplified PolymorphicDNA (RAPD) Markers PDFDocumento12 pagine(P) Random Amplified PolymorphicDNA (RAPD) Markers PDFDewi Kartika DekaNessuna valutazione finora
- A Single Copy of Virus-Derived, Transgene-Encoding Hairpin RNA Confers BYDV ImmunityDocumento5 pagineA Single Copy of Virus-Derived, Transgene-Encoding Hairpin RNA Confers BYDV ImmunityDewi Kartika DekaNessuna valutazione finora
- Evolution DBQDocumento4 pagineEvolution DBQCharles JordanNessuna valutazione finora
- SKF CMSS2200 PDFDocumento2 pagineSKF CMSS2200 PDFSANTIAGONessuna valutazione finora
- DYCONEX Materials en 2020 ScreenDocumento2 pagineDYCONEX Materials en 2020 ScreenhhaiddaoNessuna valutazione finora
- Vectors Notes (Answers)Documento24 pagineVectors Notes (Answers)ScionNessuna valutazione finora
- The Neuroscience of Helmholtz and The Theories of Johannes Muèller Part 2: Sensation and PerceptionDocumento22 pagineThe Neuroscience of Helmholtz and The Theories of Johannes Muèller Part 2: Sensation and PerceptionCrystal JenningsNessuna valutazione finora
- Validación Española ADHD-RSDocumento7 pagineValidación Española ADHD-RSCristina Andreu NicuesaNessuna valutazione finora
- DSynchronize (ENG)Documento3 pagineDSynchronize (ENG)Rekha Rajarajan100% (1)
- BS 07533-3-1997Documento21 pagineBS 07533-3-1997Ali RayyaNessuna valutazione finora
- Creative LibrarianDocumento13 pagineCreative LibrarianulorNessuna valutazione finora
- Citroen CX Manual Series 2 PDFDocumento646 pagineCitroen CX Manual Series 2 PDFFilipe Alberto Magalhaes0% (1)
- Puratattva No 41Documento3 paginePuratattva No 41ultimategoonNessuna valutazione finora
- Workplace Risk Assessment PDFDocumento14 pagineWorkplace Risk Assessment PDFSyarul NizamzNessuna valutazione finora
- Chapter 2 Axial and Torsional ElementsDocumento57 pagineChapter 2 Axial and Torsional ElementsAhmad FaidhiNessuna valutazione finora
- Help SIMARIS Project 3.1 enDocumento61 pagineHelp SIMARIS Project 3.1 enVictor VignolaNessuna valutazione finora
- Feasibility and Optimization of Dissimilar Laser Welding ComponentsDocumento366 pagineFeasibility and Optimization of Dissimilar Laser Welding Componentskaliappan45490Nessuna valutazione finora
- Machine Design REE 302: CH 1: Introduction To Mechanical Engineering DesignDocumento26 pagineMachine Design REE 302: CH 1: Introduction To Mechanical Engineering DesignDull PersonNessuna valutazione finora
- Graduate Macro Theory II: The Real Business Cycle Model: Eric Sims University of Notre Dame Spring 2017Documento25 pagineGraduate Macro Theory II: The Real Business Cycle Model: Eric Sims University of Notre Dame Spring 2017Joab Dan Valdivia CoriaNessuna valutazione finora
- Data Sheet: Item N°: Curve Tolerance According To ISO 9906Documento3 pagineData Sheet: Item N°: Curve Tolerance According To ISO 9906Aan AndianaNessuna valutazione finora
- Grimm (2015) WisdomDocumento17 pagineGrimm (2015) WisdomBruce WayneNessuna valutazione finora
- Distillation Column DesignDocumento42 pagineDistillation Column DesignAakanksha Raul100% (1)
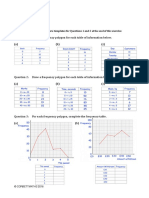
- Frequency Polygons pdf3Documento7 pagineFrequency Polygons pdf3Nevine El shendidyNessuna valutazione finora
- Surge Arrester PresentationDocumento63 pagineSurge Arrester PresentationRamiro FelicianoNessuna valutazione finora
- Curriculum Improvement v2Documento47 pagineCurriculum Improvement v2Nica Lagrimas100% (1)
- Analysis and Design of Well FoundationDocumento40 pagineAnalysis and Design of Well Foundationdhanabal100% (1)
- The Adoption of e Procurement in Tanzani PDFDocumento5 pagineThe Adoption of e Procurement in Tanzani PDFDangyi GodSeesNessuna valutazione finora
- Handbook+for+Participants+ +GCC+TeenDocumento59 pagineHandbook+for+Participants+ +GCC+Teenchloe.2021164Nessuna valutazione finora
- Slipform Construction TechniqueDocumento6 pagineSlipform Construction TechniqueDivyansh NandwaniNessuna valutazione finora
- Question BankDocumento3 pagineQuestion BankHimanshu SharmaNessuna valutazione finora
- Native Instruments Sibelius Sound Sets - The Sound Set ProjectDocumento3 pagineNative Instruments Sibelius Sound Sets - The Sound Set ProjectNicolas P.Nessuna valutazione finora
- Slip Ring Motor CheckDocumento3 pagineSlip Ring Motor CheckRolando LoayzaNessuna valutazione finora