Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Assignment of Resiration SyetemDocumento5 pagineAssignment of Resiration SyetemarsalanbookNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Ambulance Medical EquipmentDocumento32 pagineAmbulance Medical EquipmentarsalanbookNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- 1000 Most Common Words (SAT)Documento70 pagine1000 Most Common Words (SAT)grellian95% (20)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
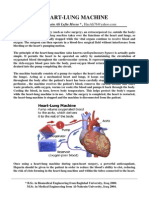
- Heart-Lung MachineDocumento4 pagineHeart-Lung Machinearsalanbook100% (2)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- European Society of Cardiology Congress - Augast.2022Documento36 pagineEuropean Society of Cardiology Congress - Augast.2022Giorgi PopiashviliNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- 2009 Handbook of Nurse Anesthesia, John J. Nagelhout, Karen Plaus PDFDocumento840 pagine2009 Handbook of Nurse Anesthesia, John J. Nagelhout, Karen Plaus PDFNoval Zain100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Clinical Indications, Treatment and Current PracticeDocumento14 pagineClinical Indications, Treatment and Current PracticefadmayulianiNessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Unit1Documento40 pagineUnit1Lily KkNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- Nayeli Sierra Ga FinalDocumento7 pagineNayeli Sierra Ga Finalapi-300744043Nessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- Beginning Medical Transcription (2nd Ed.) : Cardiopulmonary Reports Ent and Ophthalmology ReportsDocumento7 pagineBeginning Medical Transcription (2nd Ed.) : Cardiopulmonary Reports Ent and Ophthalmology ReportsRoderick RichardNessuna valutazione finora
- Diabetic DysipidemiaDocumento8 pagineDiabetic DysipidemiaAyur SuccessNessuna valutazione finora
- Heat Therapy To Reduce Chest Pain Among Patient With ACSDocumento10 pagineHeat Therapy To Reduce Chest Pain Among Patient With ACSRavitman RavitmanNessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Top 10 Ways to Lower Your Triglycerides NaturallyDocumento2 pagineTop 10 Ways to Lower Your Triglycerides NaturallySumit DhingraNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Mapeh 7-With TosDocumento5 pagineMapeh 7-With TosLorraine Calvez Donio100% (1)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- ExerciseDocumento7 pagineExerciseAjay JethwaNessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Medicinal Flora in Holy QuranDocumento16 pagineMedicinal Flora in Holy QuranMuhammad Zakiy FadlullahNessuna valutazione finora
- A Study To Assess The Knowledge About Risk Factors and Warning Signs of Acute Coronary Syndrome Among Patients Admitted in Cardiac Medical Unit at Sctimst, Triv AndrumDocumento72 pagineA Study To Assess The Knowledge About Risk Factors and Warning Signs of Acute Coronary Syndrome Among Patients Admitted in Cardiac Medical Unit at Sctimst, Triv AndrumNise Mon KuriakoseNessuna valutazione finora
- 4e 2ndproof ch7 PDFDocumento19 pagine4e 2ndproof ch7 PDFJessa AquitanNessuna valutazione finora
- Medical-Surgical Management For RHDDocumento4 pagineMedical-Surgical Management For RHDJansen Arquilita RiveraNessuna valutazione finora
- Hypertension: Management and TreatmentDocumento9 pagineHypertension: Management and TreatmentSheila May Teope SantosNessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Cardiovascular NotebookDocumento140 pagineCardiovascular NotebookChantal CarnesNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- Final Announcement ACIC ACEM SCU CME PDFDocumento23 pagineFinal Announcement ACIC ACEM SCU CME PDFFredik JackyNessuna valutazione finora
- ACC Handbook Ascvd Type 2 Diabetes: On andDocumento10 pagineACC Handbook Ascvd Type 2 Diabetes: On andZH. omg sarNessuna valutazione finora
- DLP-Bohol - Science9 Q1 W2 D4Documento6 pagineDLP-Bohol - Science9 Q1 W2 D4Marfe MontelibanoNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Completely Reverse Heart Disease With This Revolutionary TreatmentDocumento8 pagineCompletely Reverse Heart Disease With This Revolutionary TreatmentSamee's Music100% (3)
- Cardiac RehabitationDocumento4 pagineCardiac Rehabitationsundar_kumar0Nessuna valutazione finora
- 16 Vascular Diseases of Nervous System-QDocumento24 pagine16 Vascular Diseases of Nervous System-QAdi PomeranzNessuna valutazione finora
- Case Stydy Angina PectorisDocumento46 pagineCase Stydy Angina PectorissharenNessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Glenn R Gibson - Food Science and Technology Bulletin - Functional Foods. Vol. 1 PDFDocumento107 pagineGlenn R Gibson - Food Science and Technology Bulletin - Functional Foods. Vol. 1 PDFguiovanaNessuna valutazione finora
- 2000 Bookmatter TextbookOfAngiologyDocumento19 pagine2000 Bookmatter TextbookOfAngiologydavidmarkovic032Nessuna valutazione finora
- Stress and Immune SystemDocumento15 pagineStress and Immune SystemRhishabhNessuna valutazione finora
- Health Benefits of Wheat and Its Nutritional ValueDocumento13 pagineHealth Benefits of Wheat and Its Nutritional ValueJaime BallesterosNessuna valutazione finora
- What Is Coronary Angiography?Documento4 pagineWhat Is Coronary Angiography?emman_abzNessuna valutazione finora
- Health Combinat 2Documento58 pagineHealth Combinat 2mariuscornelNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)