Potrebbero piacerti anche
- Biologia - ResumosDocumento10 pagineBiologia - ResumosFrancisca VenturaNessuna valutazione finora
- Clique Aqui para Realizar A Atividade de Estudo 01 - Prazo Final - 18-06-2023 - Revisão Da TentativaDocumento8 pagineClique Aqui para Realizar A Atividade de Estudo 01 - Prazo Final - 18-06-2023 - Revisão Da TentativaJesse Santana Costa100% (1)
- 200 Questões Fisiologia HumanaDocumento37 pagine200 Questões Fisiologia HumanaDayanne Duarte Barros100% (4)
- Retículo endoplasmático e ribossomos na biologia celularDocumento13 pagineRetículo endoplasmático e ribossomos na biologia celularJuliana LeiteNessuna valutazione finora
- Biologia FT 4 - Síntese Proteica e Mitose - Exerc Cicios de ExameDocumento7 pagineBiologia FT 4 - Síntese Proteica e Mitose - Exerc Cicios de ExamemalcinacmatiasNessuna valutazione finora
- Casos Clinicos FeitosDocumento10 pagineCasos Clinicos FeitosleolinafranklinNessuna valutazione finora
- Funções organelas citologiaDocumento2 pagineFunções organelas citologiaLorena B0% (1)
- C1 CursoE Biologia 20aulasDocumento24 pagineC1 CursoE Biologia 20aulasHancock8683% (6)
- A-6 Prova AngloDocumento16 pagineA-6 Prova AngloLucas Bernardo62% (13)
- Introdução Ao Estudo Da CélulaDocumento25 pagineIntrodução Ao Estudo Da Céluladrifrancas88100% (10)
- Aula de FeridasDocumento91 pagineAula de FeridasleolinafranklinNessuna valutazione finora
- Resumos Biologia Geologia (BIOLOGIA) 11º AnoDocumento69 pagineResumos Biologia Geologia (BIOLOGIA) 11º AnoVítor Perfeito100% (1)
- Caso Clinico LindalvaDocumento4 pagineCaso Clinico LindalvaleolinafranklinNessuna valutazione finora
- Escala EstagioDocumento1 paginaEscala EstagioleolinafranklinNessuna valutazione finora
- Caso Clinico LindalvaDocumento4 pagineCaso Clinico LindalvaleolinafranklinNessuna valutazione finora
- Reunião CCIH do Hospital São Vicente FerrerDocumento1 paginaReunião CCIH do Hospital São Vicente Ferrerleolinafranklin75% (4)
- Atividade de FisicaDocumento1 paginaAtividade de FisicaleolinafranklinNessuna valutazione finora
- IVAS e Síndrome de CrupeDocumento90 pagineIVAS e Síndrome de CrupeleolinafranklinNessuna valutazione finora
- FisiologiaDocumento1 paginaFisiologialeolinafranklinNessuna valutazione finora
- Cronograma Modulo Agressão e Defesa p3 Unidade II 08 09 NDocumento3 pagineCronograma Modulo Agressão e Defesa p3 Unidade II 08 09 NleolinafranklinNessuna valutazione finora
- AmamentaçãoDocumento1 paginaAmamentaçãoleolinafranklinNessuna valutazione finora
- 2 Ano RecuperaçãoDocumento1 pagina2 Ano RecuperaçãoleolinafranklinNessuna valutazione finora
- Aldeidos e CetonasDocumento25 pagineAldeidos e CetonasleolinafranklinNessuna valutazione finora
- Sae 2Documento4 pagineSae 2leolinafranklinNessuna valutazione finora
- 06 03 TricomoniaseDocumento11 pagine06 03 TricomoniaseleolinafranklinNessuna valutazione finora
- Tricomonas VaginalisDocumento7 pagineTricomonas VaginalisleolinafranklinNessuna valutazione finora
- Objetivos Da Amamentação Semana Mundial 2009Documento1 paginaObjetivos Da Amamentação Semana Mundial 2009leolinafranklinNessuna valutazione finora
- AMAMENTAÇÃODocumento1 paginaAMAMENTAÇÃOleolinafranklinNessuna valutazione finora
- PopDocumento6 paginePopleolinafranklinNessuna valutazione finora
- Biossegurana 2012slidDocumento48 pagineBiossegurana 2012slidLeonardo QuissakNessuna valutazione finora
- GLICOCORTICÓIDES CompletoDocumento16 pagineGLICOCORTICÓIDES CompletoleolinafranklinNessuna valutazione finora
- Solicitação de LeitosDocumento1 paginaSolicitação de LeitosleolinafranklinNessuna valutazione finora
- Humanização Da AssistênciaDocumento15 pagineHumanização Da AssistêncialeolinafranklinNessuna valutazione finora
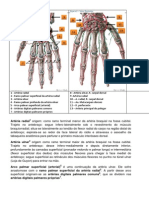
- Anatomia Irrigação Da Mão Leolina e MakleneDocumento2 pagineAnatomia Irrigação Da Mão Leolina e MakleneleolinafranklinNessuna valutazione finora
- Genetic ADocumento13 pagineGenetic AleolinafranklinNessuna valutazione finora
- AconselhamentoDocumento1 paginaAconselhamentoleolinafranklinNessuna valutazione finora
- Saúde Da MulherDocumento3 pagineSaúde Da MulherleolinafranklinNessuna valutazione finora
- Elementos PréDocumento1 paginaElementos PréleolinafranklinNessuna valutazione finora
- Obesidade, SM e DM-Tipo 2Documento47 pagineObesidade, SM e DM-Tipo 2leolinafranklin100% (1)
- Curso de Biologia Molecular Básica: Transcrição e TraduçãoDocumento22 pagineCurso de Biologia Molecular Básica: Transcrição e TraduçãoCamila MacielNessuna valutazione finora
- Sebenta BBCDocumento24 pagineSebenta BBCRita ReisNessuna valutazione finora
- Genética e Leis de Mendel: estrutura do DNA e RNADocumento41 pagineGenética e Leis de Mendel: estrutura do DNA e RNACélia FigueiredoNessuna valutazione finora
- Files AP. 1 Biologia 1Documento25 pagineFiles AP. 1 Biologia 1talles123Nessuna valutazione finora
- GLEDSON MARTINS FAUSTINO - APC 2 Junho 1ºanoDocumento2 pagineGLEDSON MARTINS FAUSTINO - APC 2 Junho 1ºanoGLEDSON MARTINS FAUSTINONessuna valutazione finora
- Perguntas e respostas sobre biologia celular e molecularDocumento14 paginePerguntas e respostas sobre biologia celular e molecularAnaCristinaCorreiaNessuna valutazione finora
- Lista de ExercíciosDocumento6 pagineLista de Exercíciosdi.loucaNessuna valutazione finora
- Biologia Molecular APX2Documento5 pagineBiologia Molecular APX2helenice marciaNessuna valutazione finora
- DNA e a estrutura em dupla héliceDocumento65 pagineDNA e a estrutura em dupla hélicelara felgueirasNessuna valutazione finora
- Ciclo celular e tradução eucarióticaDocumento13 pagineCiclo celular e tradução eucarióticaFernando Henrique DeslockNessuna valutazione finora
- Resumo Da Unidade 1.1 Bases Biológicas Do ComportamentoDocumento2 pagineResumo Da Unidade 1.1 Bases Biológicas Do ComportamentoAndrey MarquesNessuna valutazione finora
- Citoplasma PPTDocumento57 pagineCitoplasma PPTGleise CardosoNessuna valutazione finora
- Apostila BioinformáticaDocumento15 pagineApostila BioinformáticaLívio FigueirêdoNessuna valutazione finora
- Transporte ativo e transporte em quantidade na célulaDocumento27 pagineTransporte ativo e transporte em quantidade na célulaDavi ÁlefeNessuna valutazione finora
- Exercícios - 1 AvaliaçãoDocumento6 pagineExercícios - 1 AvaliaçãoLeo DiasNessuna valutazione finora
- Ácidos Nucléiocos DNA e RNADocumento64 pagineÁcidos Nucléiocos DNA e RNABrunoNessuna valutazione finora
- Apostila de Microbiologia BásicaDocumento62 pagineApostila de Microbiologia BásicaMayra PatríciaNessuna valutazione finora
- Exercícios de Biologia Molecular: transcrição, tradução e replicaçãoDocumento2 pagineExercícios de Biologia Molecular: transcrição, tradução e replicaçãoVanessa OliveiraNessuna valutazione finora
- Celula Unidade Basica Dos Seres VivosDocumento110 pagineCelula Unidade Basica Dos Seres VivosCarlos SilvaNessuna valutazione finora
- Aula II Citologia PDFDocumento32 pagineAula II Citologia PDFJaunilson CruzNessuna valutazione finora
- Células: estrutura e classificaçãoDocumento28 pagineCélulas: estrutura e classificaçãoLuciana0% (1)