Potrebbero piacerti anche
- Osteosarcoma, el cáncer de huesos más comúnDocumento10 pagineOsteosarcoma, el cáncer de huesos más comúnStephanie DomínguezNessuna valutazione finora
- Neuropatía DiabéticaDocumento15 pagineNeuropatía DiabéticaAlexandra DuránNessuna valutazione finora
- Farmacos Colinergicos...Documento51 pagineFarmacos Colinergicos...Peter RGNessuna valutazione finora
- ADULTO Y ANCIANO 2° ParcialDocumento18 pagineADULTO Y ANCIANO 2° ParcialraquelNessuna valutazione finora
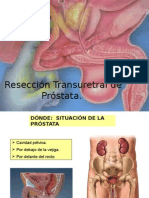
- Caso Clinico ProstataDocumento32 pagineCaso Clinico ProstataMaritza Johana Gil MartinezNessuna valutazione finora
- Atención Primaria para La SaludDocumento13 pagineAtención Primaria para La SaludVilla CelenaNessuna valutazione finora
- Epistemología de La Medicina10Documento18 pagineEpistemología de La Medicina10RichartLopezSalazarNessuna valutazione finora
- HISTORIA CLÍNICA DE TATIANA CARVAJAL CON URTICARIADocumento2 pagineHISTORIA CLÍNICA DE TATIANA CARVAJAL CON URTICARIAGabriel CarvajalNessuna valutazione finora
- Sintomas y Signos Generales de La EnfermedadDocumento7 pagineSintomas y Signos Generales de La EnfermedadAndrés Sebastián González Hidalgo100% (2)
- Semiologia RespiratoriaDocumento41 pagineSemiologia RespiratoriaAlder MirandaNessuna valutazione finora
- Especialistas en El Área de ConductaDocumento4 pagineEspecialistas en El Área de ConductaAngela RodriguezNessuna valutazione finora
- Enfermedades PulmonaresDocumento81 pagineEnfermedades PulmonaresValerie Le Noir50% (2)
- Congreso ECM - Conferencia EnfermeríaDocumento27 pagineCongreso ECM - Conferencia EnfermeríaFernando GarciaNessuna valutazione finora
- BOCIODocumento7 pagineBOCIOMarse AlvarezNessuna valutazione finora
- Caso Clinico NACDocumento44 pagineCaso Clinico NACFausto Ftr SilvaNessuna valutazione finora
- ENFERMEDADES CARENCIALES: ESQUISTOSIS Y RAQUITISMODocumento24 pagineENFERMEDADES CARENCIALES: ESQUISTOSIS Y RAQUITISMOGustavoSánchezNessuna valutazione finora
- Crioterapia, efectos y aplicaciones en fisioterapiaDocumento15 pagineCrioterapia, efectos y aplicaciones en fisioterapiaWicho HdzNessuna valutazione finora
- TesisDocumento109 pagineTesisHipradQkisNessuna valutazione finora
- Relaciones de Ciencias Sociales y La SaludDocumento12 pagineRelaciones de Ciencias Sociales y La SaludFrank Valdivia AguirreNessuna valutazione finora
- Pae - EmergenciaDocumento39 paginePae - EmergenciaRuben Antonio Taype SanchezNessuna valutazione finora
- Bacterias de Importancia MédicaDocumento7 pagineBacterias de Importancia MédicaMoisés Salazar0% (1)
- NC Envenenamiento Por AlacranDocumento6 pagineNC Envenenamiento Por AlacranPaoo Allende100% (1)
- Preeclampsia SobreagragadaDocumento4 paginePreeclampsia SobreagragadaArmando Reyes EsquincaNessuna valutazione finora
- Cancer de MamaDocumento3 pagineCancer de MamaSamuel Vicente RomeroNessuna valutazione finora
- Medicina Tradicional Peruana: Plantas y CuranderosDocumento4 pagineMedicina Tradicional Peruana: Plantas y Curanderosabel garciaNessuna valutazione finora
- Signos vitales en pediatría: frecuencia respiratoriaDocumento13 pagineSignos vitales en pediatría: frecuencia respiratoriaAdri TrejosNessuna valutazione finora
- Crecimiento de Cavidades CardiacasDocumento3 pagineCrecimiento de Cavidades CardiacasHennifer Atienza MonteroNessuna valutazione finora
- Citología Cervico UterinaDocumento15 pagineCitología Cervico UterinaRakissofiaNessuna valutazione finora
- Generalidades Del Examen FisicoDocumento48 pagineGeneralidades Del Examen FisicoJuan Guillermo Pinzón Rodríguez100% (1)
- Tema 2 Historia ClinicaDocumento3 pagineTema 2 Historia ClinicaSebastianJesusFierroFalconNessuna valutazione finora
- Semiologia - HC TEORIADocumento42 pagineSemiologia - HC TEORIARoxio Stefany Caasanova AlegriaNessuna valutazione finora
- PC I Parcial 1 Resumen Primer Departamental PCDocumento71 paginePC I Parcial 1 Resumen Primer Departamental PCFlores González AlejandraNessuna valutazione finora
- Nuevos factores de riesgo de FA en deportista con apnea del sueñoDocumento17 pagineNuevos factores de riesgo de FA en deportista con apnea del sueñoJosé Luis AñorgaNessuna valutazione finora
- Diagnóstico de acidosis y alcalosis metabólicaDocumento7 pagineDiagnóstico de acidosis y alcalosis metabólicadomeNessuna valutazione finora
- Concepto de Salud - Modulo1Documento51 pagineConcepto de Salud - Modulo1clawaissNessuna valutazione finora
- Consentimiento InformadoDocumento5 pagineConsentimiento InformadoNéstor FREE FIRENessuna valutazione finora
- Fármacos en La Insuficiencia Cardiaca CardiotónicosDocumento43 pagineFármacos en La Insuficiencia Cardiaca CardiotónicosBeltor80Nessuna valutazione finora
- Caso Clinico Distrofia Muscular de BeckerDocumento8 pagineCaso Clinico Distrofia Muscular de BeckerAlexis ToalaNessuna valutazione finora
- La relación médico-paciente: empatía, comunicación y humanismo en la práctica médicaDocumento44 pagineLa relación médico-paciente: empatía, comunicación y humanismo en la práctica médicaJhonny SayayNessuna valutazione finora
- Qué Es La Planificación FamiliarDocumento16 pagineQué Es La Planificación FamiliarNataly Grandeppieno RNessuna valutazione finora
- Saber - ComerDocumento3 pagineSaber - Comerjair mendezNessuna valutazione finora
- Perspectivas mundiales sobre trastornos de líquidos y electrolitosDocumento27 paginePerspectivas mundiales sobre trastornos de líquidos y electrolitosAndrea GuaranguayNessuna valutazione finora
- Tema 2 Alcanos PDFDocumento81 pagineTema 2 Alcanos PDFAlondra LunaNessuna valutazione finora
- Examen Físico CardiovascularDocumento21 pagineExamen Físico CardiovascularMelanii Carmen MariiaNessuna valutazione finora
- Imagenologia de AbdomenDocumento106 pagineImagenologia de AbdomenEnrique CarrilloNessuna valutazione finora
- Fuerza Vital ApunteDocumento2 pagineFuerza Vital ApunteMiranda Lara ItziaNessuna valutazione finora
- 24 Examen ReumatologíaDocumento59 pagine24 Examen Reumatologíavictoria martinezNessuna valutazione finora
- ApendicitisDocumento23 pagineApendicitisFreya HernandezNessuna valutazione finora
- Medicina en La Edad MediaDocumento3 pagineMedicina en La Edad MediaJeissuby GuzmánNessuna valutazione finora
- Metabolismo Diapositivas.Documento10 pagineMetabolismo Diapositivas.VILMA ROCIO USECHE CSTRONessuna valutazione finora
- Radiologia Del Aparato RespiratorioDocumento55 pagineRadiologia Del Aparato RespiratorioBelem Saenz RodasNessuna valutazione finora
- AntineoplasicosDocumento45 pagineAntineoplasicoskpcc18Nessuna valutazione finora
- Enfermedades Difusas Del Tejido ConjuntivoDocumento59 pagineEnfermedades Difusas Del Tejido ConjuntivoLiiLy GarciiaNessuna valutazione finora
- Fit Sumeria, Asiria, Babilonia JulioDocumento15 pagineFit Sumeria, Asiria, Babilonia JulioJulio Vicente SernaNessuna valutazione finora
- 5 Causalidad y Asociacion en Epidemiologia Clase No 5Documento24 pagine5 Causalidad y Asociacion en Epidemiologia Clase No 5Xavier DuranNessuna valutazione finora
- Examen Fisico RegionalDocumento19 pagineExamen Fisico RegionalNey Rumpelstiltskin100% (1)
- BMCII CuestionarioMetabolismoII MaderoHurtadoBrainMichelDocumento3 pagineBMCII CuestionarioMetabolismoII MaderoHurtadoBrainMichelBrain LongwoodNessuna valutazione finora
- Metabolismo de Las ProteínasDocumento40 pagineMetabolismo de Las ProteínasFrank Sánchez Estela100% (1)
- Protein AsDocumento29 pagineProtein AsGabii PachecoNessuna valutazione finora
- Metabolismo de Las ProteinasDocumento5 pagineMetabolismo de Las ProteinasBryan S CalleNessuna valutazione finora
- Mantenimiento vehículos PRLDocumento4 pagineMantenimiento vehículos PRLJorge Catalan CatalanNessuna valutazione finora
- Compendio Normativo Hig Seg NicaraguaDocumento585 pagineCompendio Normativo Hig Seg NicaraguaAnnieB100% (2)
- Ley 185, Código Del Trabajo de NicaraguaDocumento94 pagineLey 185, Código Del Trabajo de NicaraguaDereck Lyon75% (4)
- Presentacin Hot Potatoes 1203985178927491 2Documento16 paginePresentacin Hot Potatoes 1203985178927491 2Luis CarranzaNessuna valutazione finora
- DiseñoDocumento5 pagineDiseñoLuis CarranzaNessuna valutazione finora
- Clasificación de Los RobotsDocumento5 pagineClasificación de Los RobotsLuis CarranzaNessuna valutazione finora
- RequerimientosDocumento5 pagineRequerimientosLuis CarranzaNessuna valutazione finora
- Analizar El Impulso ElectricoDocumento3 pagineAnalizar El Impulso ElectricoLuis CarranzaNessuna valutazione finora
- Marco Inf UTPDocumento8 pagineMarco Inf UTPDanielSaniNessuna valutazione finora
- Núñez - Ari016 - Actividad 5Documento7 pagineNúñez - Ari016 - Actividad 5Eli DamnNessuna valutazione finora
- Carta A Una Madre o A Un Padre SobreprotectorDocumento3 pagineCarta A Una Madre o A Un Padre SobreprotectorIreri RobledoNessuna valutazione finora
- Cantonizaciond e UrdanetaDocumento3 pagineCantonizaciond e UrdanetaZulema FernandezNessuna valutazione finora
- Desague Cloacal Calculo de Pendiente3Documento10 pagineDesague Cloacal Calculo de Pendiente3muviar_Nessuna valutazione finora
- Cifras Significativas y RedondeoDocumento12 pagineCifras Significativas y RedondeoBrayan Jossep Huajalsaico CespedesNessuna valutazione finora
- Ejercicios de PuntuaciónDocumento2 pagineEjercicios de PuntuaciónAndyNessuna valutazione finora
- Física1 Grupo5 Caballero FabrizioDocumento17 pagineFísica1 Grupo5 Caballero FabrizioFabrizio Caballero VillasantiNessuna valutazione finora
- Actividad T4-01 MAPA MENTALDocumento2 pagineActividad T4-01 MAPA MENTALCarmenNessuna valutazione finora
- Artículo Olga CostaDocumento6 pagineArtículo Olga CostaVictor DuarteNessuna valutazione finora
- Primer Parcial Biociencias IIDocumento4 paginePrimer Parcial Biociencias IIJim MorrisonNessuna valutazione finora
- Leadbeater Charles - Ultimas Treinta Vidas de AlcioneDocumento285 pagineLeadbeater Charles - Ultimas Treinta Vidas de AlcioneComunidad Santimonia100% (1)
- 416268522-SESION-DE-APRENDIZAJE-5to - Proyecto de VidassDocumento3 pagine416268522-SESION-DE-APRENDIZAJE-5to - Proyecto de VidassyendraoNessuna valutazione finora
- Cultura de La Paz EquipoDocumento7 pagineCultura de La Paz EquipoAna BalderasNessuna valutazione finora
- MEMORIALDocumento1 paginaMEMORIALreyna yucraNessuna valutazione finora
- Educación democrática contra las cuerdas según Marta NussbaumDocumento2 pagineEducación democrática contra las cuerdas según Marta NussbaumLuisa SuarezNessuna valutazione finora
- Recursos para El Aprendizaje Mas AmplioDocumento2 pagineRecursos para El Aprendizaje Mas AmplioMiguel AlviarezNessuna valutazione finora
- Catalogo Ball - Español Web 2021Documento105 pagineCatalogo Ball - Español Web 2021sergio andres zamora auzzaNessuna valutazione finora
- Actividad de Vienvenida de TutoriaDocumento3 pagineActividad de Vienvenida de TutoriaNeiser Mena O̶̶̶̶̶̲̲̅̅f̶̶̶̶̶̲̲̅̅ı̶̶̶̶̶̲̲̅̅c̶̶̶̶̶̲̲̅̅ı̶̶̶̶̶̲̲̅̅a̶̶̶̶̶̲̲̅̅l̶̶̶̶̶̲̲̅̅'̶̶̶̶̶̲̲̅̅F̶̶̶̶̶̲̲̅̅bNessuna valutazione finora
- Benjamin Constant. Libertad de Los Modernos y de Los AntiguosDocumento8 pagineBenjamin Constant. Libertad de Los Modernos y de Los AntiguosValeria Denisse LozanoNessuna valutazione finora
- PosturasDocumento64 paginePosturasLuis GonzalezNessuna valutazione finora
- Teoria Eje 4Documento8 pagineTeoria Eje 4Vanessa Penaloza CabreraNessuna valutazione finora
- Resurreccion de CristoDocumento9 pagineResurreccion de CristoJohendri SuarezNessuna valutazione finora
- Cepillado de DientesDocumento2 pagineCepillado de Dientesrosa pazNessuna valutazione finora
- Conducción PersonalDocumento11 pagineConducción PersonalMarco LeivaNessuna valutazione finora
- TP 4Documento7 pagineTP 4matute0% (1)
- De Qué Hablamos Cuándo Hablamos de AmorDocumento4 pagineDe Qué Hablamos Cuándo Hablamos de AmorPaulina AlondraNessuna valutazione finora
- Arde Troya 1Documento9 pagineArde Troya 1Vanesa BiancoNessuna valutazione finora
- Profecias Parravicini y TextoDocumento170 pagineProfecias Parravicini y TextoPablo AC Trabucco100% (2)
- 1-Mapa Conceptual Actividad Semana 4Documento2 pagine1-Mapa Conceptual Actividad Semana 4FARICNessuna valutazione finora