Potrebbero piacerti anche
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- PolDocumento2 paginePolReno AndriNessuna valutazione finora
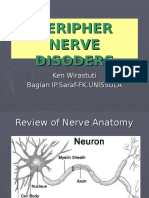
- Peripheral Nerve Disorders ReviewDocumento66 paginePeripheral Nerve Disorders ReviewReno Andri0% (1)
- KIDocumento2 pagineKIReno AndriNessuna valutazione finora
- PoloDocumento3 paginePoloReno AndriNessuna valutazione finora
- The Defense Mechanism Manual SummaryDocumento37 pagineThe Defense Mechanism Manual SummaryPriya PuriNessuna valutazione finora
- KLBDocumento3 pagineKLBReno AndriNessuna valutazione finora
- G MaxDocumento2 pagineG MaxReno AndriNessuna valutazione finora
- LomoDocumento2 pagineLomoReno AndriNessuna valutazione finora
- LBM 2 Modul Penglihatan MMMDocumento2 pagineLBM 2 Modul Penglihatan MMMReno AndriNessuna valutazione finora
- The Defense Mechanism Manual SummaryDocumento37 pagineThe Defense Mechanism Manual SummaryPriya PuriNessuna valutazione finora
- LBM 2 Modul Saraf - NSAIDsOPIOID-edit2016 PDFDocumento52 pagineLBM 2 Modul Saraf - NSAIDsOPIOID-edit2016 PDFReno AndriNessuna valutazione finora
- 1 s2.0 S2090123210000585 MainDocumento8 pagine1 s2.0 S2090123210000585 MainReno AndriNessuna valutazione finora
- Current Opinion in Neurology Volume 28 Issue 1 2015 (Doi 10.1097 - WCO.0000000000000163) Golding, John F. Gresty, Michael A. - Pathophysiology and Treatment of Motion SicknessDocumento6 pagineCurrent Opinion in Neurology Volume 28 Issue 1 2015 (Doi 10.1097 - WCO.0000000000000163) Golding, John F. Gresty, Michael A. - Pathophysiology and Treatment of Motion SicknessReno AndriNessuna valutazione finora
- Master LBM 3Documento4 pagineMaster LBM 3Reno AndriNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- A Case Study of SAMR Model in Teaching IDocumento191 pagineA Case Study of SAMR Model in Teaching IangelNessuna valutazione finora
- Genoxidil provides anti-aging benefits from around the worldDocumento2 pagineGenoxidil provides anti-aging benefits from around the worldnathy ProañoNessuna valutazione finora
- Hamsini SreeramDocumento33 pagineHamsini SreeramkalyanpavuralaNessuna valutazione finora
- 6.conversion of Amino Acids To Specialized ProductsDocumento8 pagine6.conversion of Amino Acids To Specialized Productsقتيبه خالد دحام خلفNessuna valutazione finora
- Cellgevity Product SheetDocumento3 pagineCellgevity Product SheetVictor ManalacNessuna valutazione finora
- Glutathione Intolerance? Histamine Intolerance?: Dr. Ben LynchDocumento48 pagineGlutathione Intolerance? Histamine Intolerance?: Dr. Ben Lynchfyrtenheimer100% (2)
- Res Vet Sci - 136 - 444-452 - 2021Documento29 pagineRes Vet Sci - 136 - 444-452 - 2021Phil O'Kingfisher-ShepperdNessuna valutazione finora
- Antibacterial Action 2-Bromo-2-Nitropropane-1,3-Diol (Bronopol)Documento6 pagineAntibacterial Action 2-Bromo-2-Nitropropane-1,3-Diol (Bronopol)lokesh1331Nessuna valutazione finora
- Oxid Antioxid Med Sci 201431Documento91 pagineOxid Antioxid Med Sci 201431Med AjNessuna valutazione finora
- How NAC and Glutathione Can Help To Heal Hashimoto's DiseaseDocumento4 pagineHow NAC and Glutathione Can Help To Heal Hashimoto's Diseasemightyfever2973Nessuna valutazione finora
- Acetyl Cysteine Literature Corrections MadeDocumento22 pagineAcetyl Cysteine Literature Corrections MademagimaryNessuna valutazione finora
- Toxic Responses To The LiverDocumento18 pagineToxic Responses To The LiversaiibelievesNessuna valutazione finora
- Detoxifying Cancer Causing Agents To Prevent CancerDocumento7 pagineDetoxifying Cancer Causing Agents To Prevent CancerFedoxyzNessuna valutazione finora
- Ácido AscórbicoDocumento304 pagineÁcido AscórbicoKleberCamposMirandaFilhoNessuna valutazione finora
- Sample Ch05Documento53 pagineSample Ch05mb_13_throwawayNessuna valutazione finora
- Denamarin Chewable TabletsDocumento5 pagineDenamarin Chewable Tabletsabhijit_gothoskar6039Nessuna valutazione finora
- Fix Your Period Cleanse Prep PDFDocumento4 pagineFix Your Period Cleanse Prep PDFblizzNessuna valutazione finora
- International Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakDocumento7 pagineInternational Journal of Women's Dermatology: P.E. Grimes, MD, S. Ijaz, BA, MPH, R. Nashawati, BS, MSGM, D. KwakRiefka Ananda ZulfaNessuna valutazione finora
- Mekanisme AloksanDocumento11 pagineMekanisme AloksanYolanda Octora LimbongNessuna valutazione finora
- H2SDocumento21 pagineH2SBeatrizCamposNessuna valutazione finora
- Types of Compounds Forming Glucuronides and Their ExamplesDocumento24 pagineTypes of Compounds Forming Glucuronides and Their ExamplesMLUNGISI MkhwanaziNessuna valutazione finora
- ComingClean InternetVersionSec.Documento168 pagineComingClean InternetVersionSec.Lou DregelyNessuna valutazione finora
- Ecotoxicology and Environmental SafetyDocumento8 pagineEcotoxicology and Environmental SafetyEko RaharjoNessuna valutazione finora
- Deanna Protocol Metabolic Plan For DownloadDocumento8 pagineDeanna Protocol Metabolic Plan For Downloadniknik73Nessuna valutazione finora
- GmGSTU23 Encoding A Tau Class Glutathione S-TransfDocumento14 pagineGmGSTU23 Encoding A Tau Class Glutathione S-Transfakhileshyadav3794186Nessuna valutazione finora
- Phytotoxicity and Phytoremediation Potential of Mercury in Indian Mustard and Two Ferns With Mercury Contaminated Water and Oak Ridge Soil - 9241Documento12 paginePhytotoxicity and Phytoremediation Potential of Mercury in Indian Mustard and Two Ferns With Mercury Contaminated Water and Oak Ridge Soil - 9241レテイ サマ ヒルカNessuna valutazione finora
- Carolyn M. Matthews - Nurturing Your Divine FeminineDocumento1 paginaCarolyn M. Matthews - Nurturing Your Divine FeminineJoro SongsongNessuna valutazione finora
- Aging and ImmunityDocumento14 pagineAging and ImmunitydewdropmasNessuna valutazione finora
- What Is The Difference Between Glutathione and L - GlutathioneDocumento2 pagineWhat Is The Difference Between Glutathione and L - GlutathioneAmitav Mishra0% (2)
- To Optimize Mitochondria: 22 Natural WaysDocumento26 pagineTo Optimize Mitochondria: 22 Natural Waysreyshme100% (11)