Potrebbero piacerti anche
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Multi OrgasmDocumento29 pagineMulti Orgasm4gen_7100% (2)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Speech in ActionDocumento160 pagineSpeech in ActionSplashXNessuna valutazione finora
- Trailblazer 2019Documento75 pagineTrailblazer 2019Georgiana CiaraNessuna valutazione finora
- (TCM) Foods and ElementsDocumento25 pagine(TCM) Foods and ElementsKevin FerreiraNessuna valutazione finora
- Thinking About StrategyDocumento14 pagineThinking About StrategyMehdi FaizyNessuna valutazione finora
- Artifical Light Annotated Bibliography v3Documento7 pagineArtifical Light Annotated Bibliography v3api-308065432Nessuna valutazione finora
- Circadian Rhythms in Liver Metabolism and DiseaseDocumento10 pagineCircadian Rhythms in Liver Metabolism and Diseaseapi-308065432100% (1)
- Shah Peer Review FormDocumento2 pagineShah Peer Review Formapi-308065432Nessuna valutazione finora
- Shah CommentsDocumento14 pagineShah Commentsapi-308065432Nessuna valutazione finora
- Sleep Deficiency and Deprivation Leading To Cardiovascular DiseaseDocumento5 pagineSleep Deficiency and Deprivation Leading To Cardiovascular Diseaseapi-308065432Nessuna valutazione finora
- IntroductionDocumento7 pagineIntroductionapi-308065432Nessuna valutazione finora
- The Impact of Light From Computer Monitors On Melatonin Levels in College StudentsDocumento11 pagineThe Impact of Light From Computer Monitors On Melatonin Levels in College Studentsapi-308065432Nessuna valutazione finora
- OutlineDocumento2 pagineOutlineapi-308065432Nessuna valutazione finora
- The Mammalian Circadian Timing System - Synchronization of Peripheral ClocksDocumento9 pagineThe Mammalian Circadian Timing System - Synchronization of Peripheral Clocksapi-308065432Nessuna valutazione finora
- Artifical Light Rough DraftDocumento3 pagineArtifical Light Rough Draftapi-308065432Nessuna valutazione finora
- Uncovering Residual Effects of Chronic Sleep Loss On Human PerformanceDocumento16 pagineUncovering Residual Effects of Chronic Sleep Loss On Human Performanceapi-308065432Nessuna valutazione finora
- Maternal Sleep Deprivation at Different Stages of Pregnancy Impairs The Emotional and Cognitive Functions and Suppresses Hippocampal Long-Term Potentiation in The Offspring RatsDocumento10 pagineMaternal Sleep Deprivation at Different Stages of Pregnancy Impairs The Emotional and Cognitive Functions and Suppresses Hippocampal Long-Term Potentiation in The Offspring Ratsapi-308065432Nessuna valutazione finora
- High Sensitivity of The Human Circadian Melatonin Rhythm To Resetting by Short Wavelength LightDocumento4 pagineHigh Sensitivity of The Human Circadian Melatonin Rhythm To Resetting by Short Wavelength Lightapi-308065432Nessuna valutazione finora
- Artifical Light Rough DraftDocumento13 pagineArtifical Light Rough Draftapi-308065432Nessuna valutazione finora
- Artifical Light Annotated BibliographyDocumento5 pagineArtifical Light Annotated Bibliographyapi-308065432Nessuna valutazione finora
- Protecting The Melatonin Rhythm Through Circadian Healthy Light ExposureDocumento53 pagineProtecting The Melatonin Rhythm Through Circadian Healthy Light Exposureapi-308065432Nessuna valutazione finora
- Breast Cancer and Circadian Disruption From Electric Lighting in The Modern WorldDocumento22 pagineBreast Cancer and Circadian Disruption From Electric Lighting in The Modern Worldapi-308065432Nessuna valutazione finora
- Lighting For The Human Circadian Clock - Recent Research Indicates That Lighting Has Become A Public Health IssueDocumento9 pagineLighting For The Human Circadian Clock - Recent Research Indicates That Lighting Has Become A Public Health Issueapi-308065432Nessuna valutazione finora
- Circadian Rhythms and Risk For Substance Use Disorders in AdolescenceDocumento13 pagineCircadian Rhythms and Risk For Substance Use Disorders in Adolescenceapi-308065432Nessuna valutazione finora
- The Role of Retinal Photoreceptors in The Regulation of Circadian RhythmsDocumento13 pagineThe Role of Retinal Photoreceptors in The Regulation of Circadian Rhythmsapi-308065432Nessuna valutazione finora
- OutlineDocumento2 pagineOutlineapi-308065432Nessuna valutazione finora
- Primary Paper PresentationDocumento7 paginePrimary Paper Presentationapi-308065432Nessuna valutazione finora
- Tbi Neurodegenerative Disease Annotated BibliographyDocumento2 pagineTbi Neurodegenerative Disease Annotated Bibliographyapi-308065432Nessuna valutazione finora
- Artifical Light Annotated BibliographyDocumento2 pagineArtifical Light Annotated Bibliographyapi-308065432Nessuna valutazione finora
- Sensitivity of The Human Circadian Pacemaker To Nocturnal Light - Melatonin Phase Resetting and SuppressionDocumento8 pagineSensitivity of The Human Circadian Pacemaker To Nocturnal Light - Melatonin Phase Resetting and Suppressionapi-308065432Nessuna valutazione finora
- Exposure To Room Light Before Bedtime Suppresses Melatonin Onset and Shortens Melatonin Duration in HumansDocumento10 pagineExposure To Room Light Before Bedtime Suppresses Melatonin Onset and Shortens Melatonin Duration in Humansapi-308065432Nessuna valutazione finora
- Histology AssignmentDocumento4 pagineHistology AssignmentNico LokoNessuna valutazione finora
- The Wolf and The LambDocumento4 pagineThe Wolf and The LambRaff Habibie RizzkhanbjmNessuna valutazione finora
- Science Level 4 - Animal CardsDocumento2 pagineScience Level 4 - Animal CardsClaudiaCarolinadeZeledónNessuna valutazione finora
- Kaylee Howlett ResumeDocumento2 pagineKaylee Howlett Resumeapi-353201001Nessuna valutazione finora
- Asexual Reproduction Worksheet: Name: . DateDocumento3 pagineAsexual Reproduction Worksheet: Name: . DateTamondong PaulNessuna valutazione finora
- NG TubeDocumento3 pagineNG TubeKenny JosefNessuna valutazione finora
- Dani's Odyssey or Some Commotion in The Hinterland FogDocumento228 pagineDani's Odyssey or Some Commotion in The Hinterland FogHanged Man PublishingNessuna valutazione finora
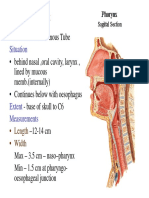
- H & N - Pharynx& Deglutition PDFDocumento17 pagineH & N - Pharynx& Deglutition PDFRamin AmirkhiziNessuna valutazione finora
- Plitvice Lakes, CroatiaDocumento7 paginePlitvice Lakes, CroatiaMirkoNessuna valutazione finora
- Imo 1.13 Elemelementary First AidDocumento22 pagineImo 1.13 Elemelementary First AidMuhammad TaufiqNessuna valutazione finora
- A Detailed Lesson Plan in Partial Fulfillment of The Course Requirements in (PED8)Documento11 pagineA Detailed Lesson Plan in Partial Fulfillment of The Course Requirements in (PED8)Angel Clarence BasilioNessuna valutazione finora
- Idoneth Deepkin Akhelian Thrallmaster v2Documento10 pagineIdoneth Deepkin Akhelian Thrallmaster v2Cerdeira HansNessuna valutazione finora
- Institute nephro-urology rate listDocumento9 pagineInstitute nephro-urology rate listVertika VermaNessuna valutazione finora
- Spiral FlapDocumento20 pagineSpiral FlapDiolanda De AbreuNessuna valutazione finora
- Food ChainsDocumento14 pagineFood ChainsShemaiah HoyteNessuna valutazione finora
- Catclass NotesDocumento3 pagineCatclass NotesIssac CaldwellNessuna valutazione finora
- ErizoDocumento36 pagineErizoEloy De Jesús SalazarNessuna valutazione finora
- Medical Parasitology: Human ParasitesDocumento30 pagineMedical Parasitology: Human ParasitesElycaNessuna valutazione finora
- Crochet Plush Bunny Neon Amigurumi Free PatternDocumento4 pagineCrochet Plush Bunny Neon Amigurumi Free PatternflechedestructriceNessuna valutazione finora
- Economically Feasible Fish Feed For GIFT Tilapia (Oreochromis LankaDocumento31 pagineEconomically Feasible Fish Feed For GIFT Tilapia (Oreochromis Lankasantosh kumarNessuna valutazione finora
- Cryo Fructo Qual KitDocumento3 pagineCryo Fructo Qual KitSuryakant HayatnagarkarNessuna valutazione finora
- Ex Situ Conservation: R.R.Bharathi 2 Yr BiochemistryDocumento20 pagineEx Situ Conservation: R.R.Bharathi 2 Yr BiochemistryVishal TakerNessuna valutazione finora
- The Market Square Dog: Name: ClassDocumento8 pagineThe Market Square Dog: Name: ClassAlejandra GonzálezNessuna valutazione finora
- Advanced Ocean Animals Activity BookDocumento5 pagineAdvanced Ocean Animals Activity BookRetno MaharaniNessuna valutazione finora
- Soal Vocabulary Part 2Documento3 pagineSoal Vocabulary Part 2Sastrawan PelawiNessuna valutazione finora