Potrebbero piacerti anche
- Moléculas de Relación CelularDocumento15 pagineMoléculas de Relación CelularAlejandro SuaresNessuna valutazione finora
- Biología Seminario Semana 6 UsmpDocumento7 pagineBiología Seminario Semana 6 Usmpdavid taqu1reNessuna valutazione finora
- Tarea S13-Grupo 4a..Documento8 pagineTarea S13-Grupo 4a..JOB PPERSNessuna valutazione finora
- Recepción CelularDocumento11 pagineRecepción CelularStephano P Carranza75% (4)
- Seminario Fisica Semana 13 BIOELECTRICIDADDocumento16 pagineSeminario Fisica Semana 13 BIOELECTRICIDADJoaquin Alcantara De la TorreNessuna valutazione finora
- Informe S5 - GRUPO 8ADocumento6 pagineInforme S5 - GRUPO 8AValeria León FallaqueNessuna valutazione finora
- Informe Fisica 9Documento10 pagineInforme Fisica 9Angelina100% (1)
- Biologia SEMANA 13Documento3 pagineBiologia SEMANA 13john salchichon100% (5)
- Informe S6 - GRUPO 8ADocumento5 pagineInforme S6 - GRUPO 8AValeria León Fallaque0% (1)
- De Gen A ProteinaDocumento2 pagineDe Gen A ProteinaDaniela Romero Rios100% (3)
- Citoplazma, Citoesqueleto y Matriz ExtracelularDocumento10 pagineCitoplazma, Citoesqueleto y Matriz ExtracelularSofia Arias-StellaNessuna valutazione finora
- Tarea 11 Fisica BioelectrecidadDocumento6 pagineTarea 11 Fisica BioelectrecidadCeleste VásquezNessuna valutazione finora
- Bio Sem InformeDocumento15 pagineBio Sem InformeXiomara Janne Linares SaldañaNessuna valutazione finora
- BIOLOGÍA SEMINARIO - USMP (Semana 1-12) - 2018Documento123 pagineBIOLOGÍA SEMINARIO - USMP (Semana 1-12) - 2018Stephano P CarranzaNessuna valutazione finora
- Densidad Relativa de Un Organo y Suero de La Sangre de UnDocumento6 pagineDensidad Relativa de Un Organo y Suero de La Sangre de UnMaria Bautista50% (2)
- Sseminario 9 Recepción CelularDocumento1 paginaSseminario 9 Recepción CelularAnonymous hkjp4qMVRsNessuna valutazione finora
- Sesión 14Documento7 pagineSesión 14Devora GilNessuna valutazione finora
- El Origen de La Vida Terrestre ModernaDocumento6 pagineEl Origen de La Vida Terrestre ModernaEnrique Moncada MapelliNessuna valutazione finora
- INFORME S10 - GRUPO Jquirozv-01Documento10 pagineINFORME S10 - GRUPO Jquirozv-01Sadae BD50% (2)
- INFORME #09 - GRUPO 36 - BIOELECTRICIDAD - DoxxDocumento8 pagineINFORME #09 - GRUPO 36 - BIOELECTRICIDAD - DoxxJosé mario Vázquez poloNessuna valutazione finora
- Informática SEM-Informe S9-Collantes-GRUPO24Documento7 pagineInformática SEM-Informe S9-Collantes-GRUPO24franco mamani rodriguezNessuna valutazione finora
- Lab Oratorio 5Documento20 pagineLab Oratorio 5api-381307464% (11)
- Tarea 6 - Grupo 21B - Bioquímica SeminarioDocumento12 pagineTarea 6 - Grupo 21B - Bioquímica SeminarioGuianela Polo100% (1)
- Informe S6.BIO - SEM-Grupo08Documento10 pagineInforme S6.BIO - SEM-Grupo08Kenner Poult Santisteban MuñozNessuna valutazione finora
- Biomecánica y Biodinámica EXPO COMPLETODocumento16 pagineBiomecánica y Biodinámica EXPO COMPLETOJosé mario Vázquez poloNessuna valutazione finora
- Informe 1 SeminarioDocumento5 pagineInforme 1 SeminarioIsaac Loyola100% (1)
- Tarea 1 Histología 03T22Documento13 pagineTarea 1 Histología 03T22Jenny TarrilloNessuna valutazione finora
- Bioelectricidad UsmpDocumento42 pagineBioelectricidad UsmpedgarNessuna valutazione finora
- Bio Prac AyudaaaDocumento2 pagineBio Prac AyudaaaXiomara Janne Linares SaldañaNessuna valutazione finora
- 09 Recepción Celular - Mecanismos MolecularesDocumento22 pagine09 Recepción Celular - Mecanismos MolecularesJunior Forttini50% (2)
- Informe 2Documento3 pagineInforme 2Milene QNessuna valutazione finora
- Pregunta 2 y 3Documento4 paginePregunta 2 y 3Kenner Poult Santisteban Muñoz100% (1)
- Informe S12 BIOSEMDocumento5 pagineInforme S12 BIOSEMAlejandro SuaresNessuna valutazione finora
- Fuerza Muscular en Una PersonaDocumento12 pagineFuerza Muscular en Una Personaapi-380297388% (8)
- Informe 12 Histología SeminarioDocumento10 pagineInforme 12 Histología SeminarioEmery Sofía C. D. BustamanteNessuna valutazione finora
- Informe S11 - Anatomia Practica - UsmpDocumento14 pagineInforme S11 - Anatomia Practica - UsmpPaola De la cruz GalánNessuna valutazione finora
- Fisica Practica MariliaDocumento5 pagineFisica Practica MariliaCristhian Jeanpierre Cueva Alvares0% (1)
- Anatomia Humana - Garganta Del Pie y PieDocumento26 pagineAnatomia Humana - Garganta Del Pie y PieFrancesco Aguirre100% (1)
- Biologia SEMANA 14Documento4 pagineBiologia SEMANA 14john salchichon86% (21)
- Informe S11 Salud Pública Seminario - Reluz 22Documento5 pagineInforme S11 Salud Pública Seminario - Reluz 22Xiomara Janne Linares SaldañaNessuna valutazione finora
- Práctica+ ElectriDocumento21 paginePráctica+ ElectriJose Jhon Suni Aslla50% (4)
- Transporte IntracelularDocumento7 pagineTransporte IntracelularRamiro Santamaria100% (1)
- Biología C.M Seminario - Informe S3.Grupo 05Documento7 pagineBiología C.M Seminario - Informe S3.Grupo 05Edwin GonzalesNessuna valutazione finora
- Exposición S8 - Moléculas de Relación CelularDocumento17 pagineExposición S8 - Moléculas de Relación CelularAlejandro SuaresNessuna valutazione finora
- Determinacion de La Fuerza Muscular de Una PersonaDocumento1 paginaDeterminacion de La Fuerza Muscular de Una PersonaChristian Raul Rojas TarrilloNessuna valutazione finora
- Informe S8 - INFORMÁTICA I GRUPO 8Documento7 pagineInforme S8 - INFORMÁTICA I GRUPO 8José mario Vázquez poloNessuna valutazione finora
- Biologia SEMANA 15Documento6 pagineBiologia SEMANA 15john salchichon100% (6)
- Informe 3 Histología PrácticaDocumento9 pagineInforme 3 Histología PrácticaEmery Sofía C. D. BustamanteNessuna valutazione finora
- Tolerancia Inmunológica en El EmbarazoDocumento10 pagineTolerancia Inmunológica en El EmbarazoRichard Rivera0% (1)
- Semana 7 - Práctica-Ciclosis y Observación de OrganelasDocumento10 pagineSemana 7 - Práctica-Ciclosis y Observación de OrganelasjuanNessuna valutazione finora
- Tarea S12 - Grupo 4aDocumento6 pagineTarea S12 - Grupo 4awilliam david lozano gaonaNessuna valutazione finora
- Moléculas de Relación CelularDocumento16 pagineMoléculas de Relación Celularrenzo reyes rodriguez50% (2)
- Informe S5 (Presión Manométrica Pulmonar)Documento10 pagineInforme S5 (Presión Manométrica Pulmonar)Noelia Rojas100% (1)
- Hidrostática I y IIDocumento5 pagineHidrostática I y IICelesteNessuna valutazione finora
- Informe Física Pract s8Documento5 pagineInforme Física Pract s8Daniela RojasNessuna valutazione finora
- Exposición 13-Visión NormalDocumento4 pagineExposición 13-Visión NormalCeleste VásquezNessuna valutazione finora
- Informe 4 Embriología Práctica PDFDocumento12 pagineInforme 4 Embriología Práctica PDFEmery Sofía C. D. BustamanteNessuna valutazione finora
- Informe 6Documento7 pagineInforme 6Juliette Rosales33% (3)
- Informe 12Documento10 pagineInforme 12ChristianPLAYNessuna valutazione finora
- Tema 7 Uniones CelularesDocumento6 pagineTema 7 Uniones CelularesDavid Molina LópezNessuna valutazione finora
- Medicina TradicionalDocumento27 pagineMedicina Tradicionaljohn salchichon100% (1)
- Pasos Cortos Usmp Fisica-Biologia-Salud PublicaDocumento80 paginePasos Cortos Usmp Fisica-Biologia-Salud Publicajohn salchichon100% (5)
- Hidrostática 1Documento73 pagineHidrostática 1john salchichonNessuna valutazione finora
- Biologia SEMANA 15Documento6 pagineBiologia SEMANA 15john salchichon100% (6)
- Biologia SEMANA 14Documento4 pagineBiologia SEMANA 14john salchichon86% (21)
- Biologia SEMANA 13Documento3 pagineBiologia SEMANA 13john salchichon100% (5)
- Biologia SEMANA 12Documento3 pagineBiologia SEMANA 12john salchichon100% (2)
- Campaña AntiplagioDocumento9 pagineCampaña Antiplagiojohn salchichonNessuna valutazione finora
- Biologia SEMANA 9Documento4 pagineBiologia SEMANA 9john salchichon67% (9)
- Biologia SEMANA 7Documento2 pagineBiologia SEMANA 7john salchichon86% (7)
- Analisis de Situacion de Salud 2014 Gustavo LanattaDocumento5 pagineAnalisis de Situacion de Salud 2014 Gustavo Lanattajohn salchichonNessuna valutazione finora
- Semana4 Tejido Epitelial PDFDocumento12 pagineSemana4 Tejido Epitelial PDFInes Meza100% (1)
- Guía Tejido Epitelial 2Documento7 pagineGuía Tejido Epitelial 2Edwin La HozNessuna valutazione finora
- Radiologia de Quistes de Los MaxilaresDocumento9 pagineRadiologia de Quistes de Los MaxilaresLeyla Naomy Huamán FrancoNessuna valutazione finora
- Tumores OdontogenicosDocumento20 pagineTumores OdontogenicosErick CisnerosNessuna valutazione finora
- Histologia Practica 1Documento14 pagineHistologia Practica 1Sofía MatíasNessuna valutazione finora
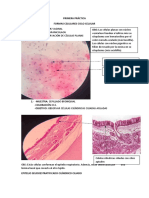
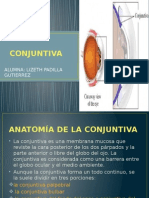
- Exposicion ConjuntivaDocumento8 pagineExposicion ConjuntivaHenry Padilla GutierrezNessuna valutazione finora
- TejidosDocumento44 pagineTejidosCARLOS EDUARDO RUIZ TAFURNessuna valutazione finora
- 02 Histología Humana - Prof. Kleider Miguel Livias Moya - Edición 2021Documento58 pagine02 Histología Humana - Prof. Kleider Miguel Livias Moya - Edición 2021KLEIDER LIVIASNessuna valutazione finora
- Elementos Celulares Epiteliales NormalesDocumento5 pagineElementos Celulares Epiteliales NormalesKristian LluayNessuna valutazione finora
- Histologia - Ojo Uabp9Documento8 pagineHistologia - Ojo Uabp9Seccion Control de Agencias URINessuna valutazione finora
- Bibliografía TejidosDocumento16 pagineBibliografía TejidosLuciana GonzálezNessuna valutazione finora
- Guia de Laboratorio Morfología Ii (MBA-151) : Tecnología MédicaDocumento45 pagineGuia de Laboratorio Morfología Ii (MBA-151) : Tecnología MédicaMiguelito MoralitoNessuna valutazione finora
- Trabajo Práctico Luciano PeraltaDocumento5 pagineTrabajo Práctico Luciano PeraltaLucianoNessuna valutazione finora
- Apunte Perna DERMATO OKDocumento43 pagineApunte Perna DERMATO OKYamila MazaNessuna valutazione finora
- HistologiaaaDocumento38 pagineHistologiaaaJuan Pablo CruzNessuna valutazione finora
- CP2 - Histología y Embriología Del Aparato RespiratorioDocumento21 pagineCP2 - Histología y Embriología Del Aparato RespiratorioAlfonso BeltránNessuna valutazione finora
- Vesicula SeminalDocumento3 pagineVesicula SeminalRahm KotaNessuna valutazione finora
- OdontoblastoDocumento6 pagineOdontoblastoJarly Lopez Portocarrero100% (1)
- Reporte de Los VideosDocumento7 pagineReporte de Los VideosAna AltamiranoNessuna valutazione finora
- Unidad 7 Sistema Reproductor Femenino y Masculino Emely OrtegaDocumento3 pagineUnidad 7 Sistema Reproductor Femenino y Masculino Emely Ortegamarcos ortegaNessuna valutazione finora
- Guía Práctica #7 Neoplasias MalignasDocumento6 pagineGuía Práctica #7 Neoplasias MalignasBryan CruzNessuna valutazione finora
- 1 Lesiones VerrucopapilaresDocumento39 pagine1 Lesiones Verrucopapilareskenia irahetaNessuna valutazione finora
- MOTAD Lab 5 y 6Documento12 pagineMOTAD Lab 5 y 6Brayan F.MNessuna valutazione finora
- Tejido EpitelialDocumento2 pagineTejido Epitelialcarlos darwinNessuna valutazione finora
- Aparato Digestivo Trabajo EscritoDocumento8 pagineAparato Digestivo Trabajo EscritoBeatriz RamirezNessuna valutazione finora
- Exámen HEG 12.06.2020 10.01hDocumento6 pagineExámen HEG 12.06.2020 10.01hmaria trentinNessuna valutazione finora
- Marco TeóricoDocumento3 pagineMarco TeóricoMariana ZuñigaNessuna valutazione finora
- Tejido EpilialDocumento17 pagineTejido Epilialmafe :3Nessuna valutazione finora
- Histología-Guia - Practica 02Documento10 pagineHistología-Guia - Practica 02Eliane Marreros BazauriNessuna valutazione finora
- 4-Sistema Foto e AudiorreceptorDocumento63 pagine4-Sistema Foto e AudiorreceptorRosa gonzalezNessuna valutazione finora