Potrebbero piacerti anche
- TINCIONESDocumento24 pagineTINCIONESpsicologia2009100% (1)
- Resumen Neoplasias Linfoides y Mieloides. Patología RobbinsDocumento39 pagineResumen Neoplasias Linfoides y Mieloides. Patología RobbinsAilyn ForeroNessuna valutazione finora
- Resumen Capitulo Inmunología RobbinsDocumento17 pagineResumen Capitulo Inmunología RobbinsLaura Galvis33% (3)
- Histoquimica de Los CarbohidratosDocumento13 pagineHistoquimica de Los Carbohidratosjimenezwil100% (4)
- Capítulo 3 - Resumen - RobbinsDocumento19 pagineCapítulo 3 - Resumen - RobbinsJulián Martínez50% (2)
- Dermatopatologia y TincionesDocumento75 pagineDermatopatologia y TincionesAntonio Rolon100% (1)
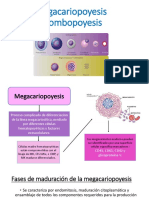
- Megacariopoyesis y TrombopoyesisDocumento5 pagineMegacariopoyesis y TrombopoyesisKatherine Chavez67% (3)
- Linfopoyesis 1 PDFDocumento97 pagineLinfopoyesis 1 PDFKim Shantal100% (2)
- Imagenes de HematologiaDocumento17 pagineImagenes de Hematologiakeiler marronNessuna valutazione finora
- Conceptos Básicos de InmunologíaDocumento7 pagineConceptos Básicos de Inmunologíaclau955100% (4)
- Alteraciones Cualitativas y Cuantitativas de La SangreDocumento26 pagineAlteraciones Cualitativas y Cuantitativas de La SangreThalía Muñoz Rivasplata92% (13)
- 3 Sem TincionesDocumento57 pagine3 Sem TincionesJosé Isaac Molina Morales100% (1)
- Hemograma de SchillingDocumento6 pagineHemograma de SchillingandreaNessuna valutazione finora
- Manual de Patología GeneralDocumento114 pagineManual de Patología GeneralAndrés Faúndez Terán100% (1)
- Reacción de ArthusDocumento6 pagineReacción de ArthusGloria María100% (2)
- Resumen. Citología y BiopsiaDocumento9 pagineResumen. Citología y BiopsiaNats Sanz Torres100% (1)
- Citología de OrinaDocumento53 pagineCitología de OrinaFrancisca50% (2)
- Cuestionario Sobre Inflamación y Reparación y Lesion CelularDocumento12 pagineCuestionario Sobre Inflamación y Reparación y Lesion CelularRoxana Salazar100% (1)
- Resumen Patologia Basica, Neoplasia, Inflamación, RegeneraciónDocumento18 pagineResumen Patologia Basica, Neoplasia, Inflamación, Regeneracióngracee275% (8)
- Hemograma de ShillingDocumento4 pagineHemograma de ShillingSilvana Quichimbo100% (1)
- Biometria HematicaDocumento92 pagineBiometria HematicaBeto Hdz100% (1)
- Automatizacion-Coagulacion 2021 IDocumento33 pagineAutomatizacion-Coagulacion 2021 IMIGUEL ANGEL TINCO QUISPENessuna valutazione finora
- LinfopoyesisDocumento19 pagineLinfopoyesisLaura Angelica de PerezNessuna valutazione finora
- Caso Clínico Inflamacion AgudaDocumento3 pagineCaso Clínico Inflamacion AgudaCarlos David Benitez0% (1)
- Sistema InmunitarioDocumento12 pagineSistema InmunitarioRigoberto Gonzales RegaladoNessuna valutazione finora
- Histologia de LaringeDocumento3 pagineHistologia de LaringeAdnan Haj Yousef Cordero100% (1)
- Enfermedad de Glanzmann (TG)Documento17 pagineEnfermedad de Glanzmann (TG)Andrea Rico DávilaNessuna valutazione finora
- Alteraciones Cuantitativas y Cualitativas de Los LeucocitosDocumento13 pagineAlteraciones Cuantitativas y Cualitativas de Los LeucocitosHector Alan Marquina Montero100% (4)
- 1-Lesión y Muerte CelularDocumento6 pagine1-Lesión y Muerte Celularapi-3727168100% (2)
- Tejido Linfoide Asociado A Bronquios (BALT)Documento6 pagineTejido Linfoide Asociado A Bronquios (BALT)Ale CaicedoNessuna valutazione finora
- Biometría HemáticaDocumento12 pagineBiometría HemáticaMarianaAcostaNessuna valutazione finora
- Procesamiento y Presentacion Del Antigeno Al Linfocito TDocumento5 pagineProcesamiento y Presentacion Del Antigeno Al Linfocito TMarciaPilcoGilNessuna valutazione finora
- TROMBOPOYESISDocumento5 pagineTROMBOPOYESISAlice0% (1)
- InterleucinasDocumento2 pagineInterleucinasJosé Damián67% (3)
- Capitulo 2 - RobbinsDocumento18 pagineCapitulo 2 - RobbinsYanina Pdv90% (10)
- Reparación de TejidosDocumento8 pagineReparación de TejidosFernando VillavicencioNessuna valutazione finora
- InmunohistoquímicaDocumento7 pagineInmunohistoquímicanachavNessuna valutazione finora
- Opsonización PDFDocumento4 pagineOpsonización PDFSelena Cedeño PicoNessuna valutazione finora
- Homograma de SchillingDocumento11 pagineHomograma de SchillingSandro OrtizNessuna valutazione finora
- Clasificación de Las Enfermedades AutoinmunesDocumento8 pagineClasificación de Las Enfermedades AutoinmunesVale Jimenez QuintanaNessuna valutazione finora
- Tejido Linfatico DifusoDocumento2 pagineTejido Linfatico Difusojulio roberto castilloNessuna valutazione finora
- Cuestionario de Patologia Cuarto Semestre FacMed ULEAMDocumento36 pagineCuestionario de Patologia Cuarto Semestre FacMed ULEAMRonald Steven Bravo Avila50% (2)
- Definición de PlaquetaDocumento4 pagineDefinición de PlaquetaThomas RamosNessuna valutazione finora
- Ebook Observaciones Del HemogramaDocumento65 pagineEbook Observaciones Del HemogramaLoli Bello SanchezNessuna valutazione finora
- LinfopoyesisDocumento12 pagineLinfopoyesisViianey Navii SpoockNessuna valutazione finora
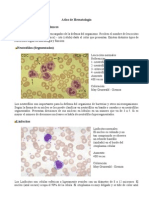
- Atlas de HematologDocumento6 pagineAtlas de Hematologanon-602585100% (2)
- Frotis de Sangre PeriféricaDocumento5 pagineFrotis de Sangre PeriféricaMarco Antonio Alonso Delgadillo100% (2)
- Pruebas de Función HepáticaDocumento18 paginePruebas de Función HepáticaJuan R. NaalNessuna valutazione finora
- Maduracion Del MonocitoDocumento3 pagineMaduracion Del MonocitoQUIMICO CLINICO WILLIANS SANCHEZ88% (8)
- Inflamación Aguda y Crónica (Preguntas)Documento10 pagineInflamación Aguda y Crónica (Preguntas)bdloedgar33% (3)
- Antiestreptolisina oDocumento7 pagineAntiestreptolisina oCristhian Jose Tejada MerinoNessuna valutazione finora
- 02 Células InmunocompetentesDocumento10 pagine02 Células InmunocompetentesIsa OcañaNessuna valutazione finora
- 2 CelulasDocumento32 pagine2 CelulasEsteban Ponce NavarroNessuna valutazione finora
- Organos LinfoidesDocumento3 pagineOrganos LinfoidesEduardo FalcónNessuna valutazione finora
- TEMA 2. Células y Órganos Del Sistema Inmune y Circulación LinfocitariaDocumento22 pagineTEMA 2. Células y Órganos Del Sistema Inmune y Circulación LinfocitariaNina Souto BlancoNessuna valutazione finora
- Resumen #2-CelulasDocumento20 pagineResumen #2-CelulasFidedigna CastroNessuna valutazione finora
- TIMODocumento4 pagineTIMOMaria Camila González100% (1)
- LINNFOPOYESISDocumento6 pagineLINNFOPOYESISChristian HidalgoNessuna valutazione finora
- TIMODocumento18 pagineTIMOangeliita1407Nessuna valutazione finora
- Timo y Educacion Timica, TareaDocumento4 pagineTimo y Educacion Timica, TareaMauricio IríasNessuna valutazione finora
- BR Realizar Estudios de Comunidad 523Documento4 pagineBR Realizar Estudios de Comunidad 523miguel angelNessuna valutazione finora
- Actividad OxidosDocumento2 pagineActividad Oxidosmiguel angelNessuna valutazione finora
- Plan de Trabajo Planilla Azul 2015Documento25 paginePlan de Trabajo Planilla Azul 2015miguel angelNessuna valutazione finora
- El Comité Encabezado Por Claudia Lizbeth Reyes Montúfar Es El Único Reconocido LegalmenteDocumento7 pagineEl Comité Encabezado Por Claudia Lizbeth Reyes Montúfar Es El Único Reconocido Legalmentemiguel angelNessuna valutazione finora
- Calendario Semestral Beta 2016 Universidad de GuanajuatoDocumento1 paginaCalendario Semestral Beta 2016 Universidad de GuanajuatoOmar Radhames UrquidezNessuna valutazione finora
- Examen Diagnostico BioquimicaDocumento4 pagineExamen Diagnostico Bioquimicamiguel angelNessuna valutazione finora
- Pronto de Visita en Tu Unidad de TrabajoDocumento5 paginePronto de Visita en Tu Unidad de Trabajomiguel angelNessuna valutazione finora
- Lista de Verificacion NOM 007 LABORATORIO HMDocumento10 pagineLista de Verificacion NOM 007 LABORATORIO HMmiguel angel100% (6)
- Analisis de BroteDocumento11 pagineAnalisis de Brotemiguel angel100% (1)
- Hymenolepis NanaDocumento11 pagineHymenolepis Nanamiguel angel50% (2)
- AnexoVIII Identificacion Recien NacidoDocumento16 pagineAnexoVIII Identificacion Recien Nacidomiguel angelNessuna valutazione finora
- Evolution of MRSADocumento9 pagineEvolution of MRSAmiguel angelNessuna valutazione finora
- Biomedicina 2010Documento98 pagineBiomedicina 2010miguel angel0% (1)
- Condiciones Sanitarias en Las Pandemias de Influenza A H1N1 de 1918 vs. 2009Documento16 pagineCondiciones Sanitarias en Las Pandemias de Influenza A H1N1 de 1918 vs. 2009miguel angelNessuna valutazione finora
- JIT Swine Flu Version 2 April 27 20091 SpanishDocumento27 pagineJIT Swine Flu Version 2 April 27 20091 Spanishmiguel angel100% (4)
- Apoptosis Vinculada A La Morfogénesis Del AstrovirusDocumento15 pagineApoptosis Vinculada A La Morfogénesis Del Astrovirusmiguel angelNessuna valutazione finora
- Libro Intraforo Uv 2008Documento185 pagineLibro Intraforo Uv 2008miguel angel0% (1)
- Vacuna Contra NeumococoDocumento22 pagineVacuna Contra Neumococomiguel angel100% (1)
- Expo Upr MaogDocumento17 pagineExpo Upr Maogmiguel angel100% (1)
- 30 Generaciones Reunidos para Celebrar El AniversarioDocumento7 pagine30 Generaciones Reunidos para Celebrar El Aniversariomiguel angelNessuna valutazione finora
- PFGEDocumento20 paginePFGEmiguel angel100% (1)
- Influenza (1)Documento17 pagineInfluenza (1)miguel angel100% (2)
- Principios Bioeticos en Terapia Genica en MéxicoDocumento15 paginePrincipios Bioeticos en Terapia Genica en Méxicomiguel angelNessuna valutazione finora
- S Aureus BasicaDocumento61 pagineS Aureus Basicamiguel angel100% (1)
- Transmision de Señales Miguel)Documento30 pagineTransmision de Señales Miguel)miguel angel100% (2)
- Transmision de Señales Miguel)Documento30 pagineTransmision de Señales Miguel)miguel angel100% (2)
- Deficiencias Del Metabolismo de Purinas y AsDocumento27 pagineDeficiencias Del Metabolismo de Purinas y Asmiguel angel100% (2)
- LPS y Receptores Toll (MAOG)Documento13 pagineLPS y Receptores Toll (MAOG)miguel angel100% (3)
- SeÑalizacion Intracelular MaogDocumento21 pagineSeÑalizacion Intracelular Maogmiguel angel100% (1)
- AminoácidosDocumento231 pagineAminoácidosJessica75% (4)
- Practica 1Documento6 paginePractica 1CINDY ELIZABETH AVILES GONZALEZNessuna valutazione finora
- 7ºTP - Ciclo CelularDocumento2 pagine7ºTP - Ciclo Celularmica turradoNessuna valutazione finora
- Examen Parcial - InmunologíaDocumento11 pagineExamen Parcial - InmunologíaJEFFERSON ALBERTO RUIZ CHAUCA100% (1)
- Hanahan Weinberg 2011.en - Es.en - Es TodoDocumento29 pagineHanahan Weinberg 2011.en - Es.en - Es TodoAlex TorresNessuna valutazione finora
- Teorico TP 10 1 BioDocumento7 pagineTeorico TP 10 1 BioCaterina Maurin 8 6 3 7 5 / 4Nessuna valutazione finora
- Ciclo de Multiplicación ViralDocumento2 pagineCiclo de Multiplicación ViralKarla OsorioNessuna valutazione finora
- Tecnicas de Tincion Del Sistema NerviosoDocumento2 pagineTecnicas de Tincion Del Sistema Nerviosoalmu19890% (1)
- Esteroides y Ejemplos BIOQUIMICADocumento33 pagineEsteroides y Ejemplos BIOQUIMICAYhara HidalgoNessuna valutazione finora
- Replicación Del DNADocumento25 pagineReplicación Del DNASayda Alisson Huacre TuctoNessuna valutazione finora
- Neurotransmisores y NeuromoduladoresDocumento21 pagineNeurotransmisores y NeuromoduladoresElizabethEsquivelCruz100% (1)
- Actividad Transporte Celular NM1 Biología - 10Documento6 pagineActividad Transporte Celular NM1 Biología - 10Ingrid ErazoNessuna valutazione finora
- Tejido Oseo y Tejdo HematopoyeticoDocumento22 pagineTejido Oseo y Tejdo Hematopoyeticojosselinmitarojas123Nessuna valutazione finora
- RIAL - Infografía Regulación MitosisDocumento1 paginaRIAL - Infografía Regulación MitosisDaviid GomezNessuna valutazione finora
- Taller Vias MetabolicasDocumento9 pagineTaller Vias MetabolicasdanielaNessuna valutazione finora
- Genes y GenomaDocumento47 pagineGenes y GenomaDiana Elam100% (1)
- Integración de Los Procesos Metabólicos. Nutri 2020Documento2 pagineIntegración de Los Procesos Metabólicos. Nutri 2020Axel LavallenNessuna valutazione finora
- FiebreDocumento28 pagineFiebreJhosbert Camacho CunoNessuna valutazione finora
- L CanavaninaDocumento6 pagineL Canavaninamo0nseerratNessuna valutazione finora
- 1 5 TejidoMuscularDocumento17 pagine1 5 TejidoMuscularOmar Sare CruzNessuna valutazione finora
- Biologia IDocumento224 pagineBiologia IEEM2DE4Nessuna valutazione finora
- SEMINARIODocumento27 pagineSEMINARIOKatherine Viviana Avila GuzmanNessuna valutazione finora
- Artículo 7Documento6 pagineArtículo 7Cele Lbo JaraNessuna valutazione finora
- Célula AnimalDocumento7 pagineCélula AnimalRolex Gonzalo100% (1)
- Guia de Aprendizaje Ciencias Naturales 8º BásicmpDocumento4 pagineGuia de Aprendizaje Ciencias Naturales 8º Básicmpkaren bascuñan100% (1)
- Protocolo #7,8 y 9Documento7 pagineProtocolo #7,8 y 9Josué Nivar De JesúsNessuna valutazione finora
- Copia de Célula VegetalDocumento33 pagineCopia de Célula VegetalLuis Felipe Perrera BarreroNessuna valutazione finora
- AminoglucósidosDocumento9 pagineAminoglucósidosBrenda0% (1)
- LipopolisacaridosDocumento14 pagineLipopolisacaridosalucard_lordhellsing100% (1)
- Espermatogenesis InfografiaDocumento2 pagineEspermatogenesis InfografiaHaruNessuna valutazione finora